ИНГИБИТОРЫ ТРАНСКРИПЦИИ И РЕПЛИКАЦИИ
А. РЕПЛИКАЦИЯ И ТРАНСКРИПЦИЯ
Синтез нуклеиновых кислот можно разделить на два этапа.
- Синтез предшественников (нуклеотидов и дезоксинуклео- тидов) из промежуточных продуктов метаболизма.
- Ферментативная полимеризация нуклеотидов с образованием макромолекул, нуклеотидные последовательности которых определяются последовательностью оснований ДНК-матрицы.
В репликации ДНК участвует много разных ферментов. Упрощенная схема этого процесса представлена на рис. 3.10. Можно отметить следующие стадии репликации.
- Цепи двойной спирали ДНК расплетаются под действием двух ферментов — ДНК-зависимой АТРазы и гиразы — и удерживаются в расплетенном состоянии благодаря связыванию со специфическим белком, называемым белком Албертса или белком 32.
- Специфическая РНК-полимераза синтезирует на каждой цепи короткие фрагменты РНК, вероятно соответствующие специфическим участкам инициации синтеза.
- Репликативная ДНК-полимераза (полимераза III уE.coli) синтезирует небольшие фрагменты ДНК (называемые фрагментами Оказаки), начиная с З'-ОН-конца РНК, используемой в качестве затравки.
- РНКаза (у Е. coli это ДНК-полимераза I, которая помимо полимеразной активности, используемой для репаративного синтеза ДНК, обладает также нуклеотидазной активностью) деградирует фрагменты РНК. ДНК-полимераза I завершает синтез фрагментов Оказаки.
- Лигаза соединяет вместе соседние фрагменты.
Процесс синтеза РНК исследован весьма тщательно. РНК- полимераза представляет собой фермент, состоящий из пяти белков: а, а', (3, р' и а. Последний белок (о) необходим для узнавания участков инициации транскрипции на ДНК- РНК- полимераза выполняет следующие функции.
- Образует комплекс с ДНК и разделяет цепи.
Фермент Е и /.ТТРаза расплетают обойную спираль Белок Ллдертса удерживает цепи в расплетенном состоянии
- Включает первый нуклеотид в правильном положении относительно ДНК (инициация).
- Включает второй нуклеотид и образует первую фосфоди- эфирную связь между первым и вторым «уклеотидами; перемещаясь вдоль молекулы ДНК, продолжает включать новые нуклеотиды и образовывать межиуклеотидные связи (элонгация).
- Останавливает процесс, когда доходит до определенной последовательности в молекуле ДНК (терминация).
f Б. ИНГИБИТОРЫ РЕПЛИКАЦИИ И ТРАНСКРИПЦИИ
Ингибиторы репликации и транскрипции можно разделить на три группы.
- Ингибиторы синтеза предшественников.
- Ингибиторы полимеризации, которые в овою очередь разделяются на а) ингибиторы матричных функций ДНК и б) ингибиторы ферментов (ДНК-реплицирующих ферментов или РНК -полимеразы).
Аналоги предшественников, которые могут нарушать как синтез предшественников, тт и полимеризацию, мы обсудим в разд. VII, посвященном антиметаболитам, а здесь рассмотрим ингибиторы матричных функций ДНК и ингибиторы полимераз (рис. 3.11).
- ИНГИБИТОРЫ МАТРИЧНЫХ ФУНКЦИЙ ДНК
Многие вещества нарушают репликацию и транскрипцию, подавляя матричные функции ДНК- Эти вещества можно разделить на две группы: 1) препараты, которые непосредственно блокируют матрицу, образуя нефункциональный комплекс с ДНК, и 2) препараты, вызывающие изменение структуры ДНК (разрывы цепей, выщепление оснований, образование ковалентных связей между двумя цепями), так что она больше не может . служить матрицей. Эти антибиотики обладают следующими свойствами.
- Связываются с ДНК самых разных клеток (бактерий, грибов или высших организмов). Поэтому их действие неспецифично, они подавляют рост любых клеток, в которые могут проникнуть, и, следовательно, очень токсичны. Они используются в медицинской практике не для лечения инфекционных заболеваний, а обычно' в качестве противоопухолевых препаратов, поскольку являются особенно активными в отношении быстрорастущих клеток.
- Подавляют синтез как РНК, так и ДНК, хотя при определенных условиях наблюдается избирательное подавление лишь одного из этих процессов.
а. Антибиотики, образующие комплексы с ДНК
al. Дауномицин (рубомицин) и адриамицин. Это антрацик- линовые антибиотики, молекула которых состоит из четырех шестиуглеродных колец, расположенных в одной плоскости. Благодаря плоской структуре такая молекула может встраиваться между парами оснований двойной спирали ДНК, стабилизируя ее и в результате этого подавляя репликацию ДНК, необходимым условием которой является расплетание цепей. Подавление
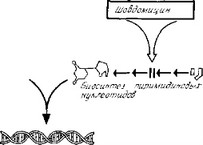
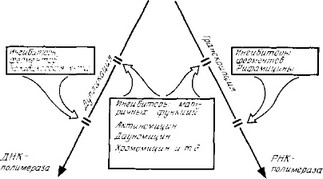
Рис. 3.11. Схема механизмов действия антибиотиков, подавляющих синтез нуклеиновых кислот. [Из работы Vergnano М., Sasselia D. (eds), Rassegna Ме- dica, Milan, Italy, Gruppo Lepetit, Spa., 1973, p. 117, с изменениями.]
синтеза РНК, вероятно, связано с тем, что молекулы антрацик- линов интеркалируют между основаниями и подавляют связывание РНК-полимеразы с ДНК. Вещества, которые действуют так же, как дауномицин, называют интеркаляторами. Дауноми- цин и его аналог адриамицин не используют для лечения инфек-
ционных заболеваний, а применяют только в химиотерапии опухолей.
а2. Актиномицин D. Это один из первых антибиотиков; он очень важен с исторической точки зрения, но слишком токсичен, чтобы его можно было использовать где-либо еще, кроме химиотерапии опухолей. Актиномицин D обратимо связывается с двухцепочечной ДНК, о чем свидетельствует изменение его спектра поглощения в видимой области в присутствии ДНК, а также изменение плавучей плотности, вязкости, кривых плавления ДНК в присутствии актиномицина. Известны два типа моделей его взаимодействия с ДНК. В моделях первого типа предполагается, что хромофор актиномицина встраивается между плоскостями оснований, как в случае дауномицина, а в моделях второго типа — что молекула антибиотика располагается в малой бороздке двойной спирали ДНК- В любом случае образование комплекса актиномицина с ДНК препятствует перемещению РНК-полимеразы вдоль ДНК-матрицы и, следовательно, подавляет синтез РНК. Подавление репликации ДНК может быть связано с тем, что ДНК в комплексе с актиномицином становится более стабильной и хуже расплетается.
аЗ. Митомицины. Эти антибиотики ковалентно связываются с ДНК, сшивая цепи молекулы и препятствуя их разделению. Благодаря природе образующейся химической связи с ДНК действие митомицинов необратимо, и эти антибиотики обладают бактерицидным действием. Очевидно, что они слишком токсичны, и их нельзя использовать в медицине в качестве антимикробных препаратов.
б. Вещества, модифицирующие ДНК
61. Блеомицины. Эти соединения образуют комплексы с ДНК, но в отличие от других ингибиторов их первичное действие, по- видимому, состоит в разрыве полинуклеотидных цепей. Блеомицины используются только в химиотерапии некоторых типов опухолевых заболеваний.
- ИНГИБИТОРЫ РНК-ПОЛИМЕРАЗЫ
Ингибиторы РНК-полимеразы распространены значительно меньше, чем ингибиторы матричных функций ДНК. Они имеют следующие общие свойства.
- Поскольку РНК-полимеразы прокариот и эукариот различаются, эти ингибиторы обычно обладают избирательным действием. Они подавляют рост или только бактериальных клеток, или только клеток животных.
- Они специфически подавляют синтез РНК в растущих бактериях и не оказывают какого-либо прямого влияния на синтез
ДНК. Остановка синтеза РНК приводит через несколько минут к остановке синтеза белка из-за отсутствия мРНК.
- Временная остановка синтеза РНК сама по себе не является гибельной для клетки. Поэтому ингибиторы РНК-полимеразы обладают бактериостатическим действием, если только их связывание с ферментом не является практически необратимым; R таком случае их действие будет бактерицидным.
а. Антибиотики — ингибиторы РНК-полимеразы
al. Анзамицины. Анзамицины — антибиотики, молекула которых включает ароматическую часть с прикрепленным к ней мостиком, представленным алифатической цепью и называемым «ручкой». Анзамицины, содержащие нафталиновое ароматическое ядро (рифамицины, стрептоварицины, толипомицины, гало- мицины), являются ингибиторами бактериальной РНК-полимеразы.
Наиболее известны среди них рифамицины, в особенности рифампин — полусинтетическое производное, широко используемое в клинике. Действие рифамицинов обусловлено практически необратимым связыванием их с РНК-полимеразой, о чем свидетельствуют 1) влияние этих антибиотиков на синтез макромолекул в целых клетках (рис. 3.12); 2) подавление in vitro бактериальной РНК-полимеразы и отсутствие эффекта в случае ДНК- полимеразы и РНК-полимеразы эукариот; 3) образование комплекса РНК-полимераза — рифамицин, который можно выделить хроматографическими методами; 4) отсутствие такого комплекса с РНК-полимеразой, полученной из бактерий, устойчивых к рифампину.
Дополнительные исследования показали, что антибиотик связывается с р-субъединицей фермента. Рифампин, очевидно, подавляет инициацию, а не элонгацию РНК, поскольку длина немногих цепей РНК, образованных в присутствии антибиотика, такая же, как и в его отсутствие. Более точно, рифампин, по-видимому, блокирует синтез РНК немедленно после образования первой фосфодиэфирной связи.
а2. Стрептолидигин. Этот антибиотик обладает слабой активностью и поэтому не используется в клинике. Стрептолидигин, как и остальные анзамицины, подавляет активность бактериальной РНК-полимеразы. Однако его механизм действия отличается от механизма действия рифамицинов, поскольку стрептолидигин подавляет и элонгацию, и инициацию синтеза РНК.
- ИНГИБИТОРЫ ДНК-ПОЛИМЕРАЗЫ
Налидиксовая кислота (Нал) подавляет репликацию ДНК в точке роста, но не подавляет репаративный синтез при рекомбинации или при залечивании радиационных повреждений. Это заключение основано на следующих наблюдениях.
- После добавления Нал к культуре чувствительных бактерий синтез ДНК останавливается раньше синтеза других макромолекул.
- Нерепликативный синтез ДНК не подавляется. Не затрагиваются такие процессы, как интеграция ДНК из донорских клеток в реципиентные клетки Е. coli при конъюгации и при трансформации В. subtilis, а также синтез ДНК после облучения Е. coli УФ-светом (даже при высоких концентрациях Нал).
- Нал не влияет на биосинтез нуклеотидов, поскольку бес- клеточная система синтеза ДНК, использующая экзогенные нуклеозидтрифосфаты, подавляется Нал.
- Недавно показано, что мишенью для Нал является гираза.
В присутствии Нал чувствительные к этому препарату клетки удлиняются, наблюдается сильная деградация ДНК и клетки погибают. Нал оказывает бактерицидное действие только на те бактерии, которые активно- синтезируют ДНК и белок. Синтез этих двух макромолекул необходим для деградации ДНК. При отмывании клеток, обработанных Нал, синтез ДНК восстанавливается, а затем восстанавливается и рост бактерий. Следовательно, Нал непрочно связывается со своей мишенью.
Относительно механизма действия двух близких антибиотиков— новобиоцина и кумермицина — сведения были весьма противоречивыми. В настоящее время, однако, установлено, что первичным действием этих антибиотиков на растущие клетки является подавление синтеза ДНК, а их мишенью является ги-
5—347
раза. Налидиксовая кислота и «овобиоцин подавляют активность двух разных субъединиц гиразы, и поэтому не наблюдается перекрестной устойчивости к этим антибиотикам.
- НЕСПЕЦИФИЧЕСКИЕ ИНГИБИТОРЫ ПОЛИМЕРАЗ
Недавно получены некоторые полусинтетические рифамици-
ны, которые, очевидно, утратили специфичность, характерную для исходных рифамицинов. Они подавляют примерно с одинаковой эффективностью функционирование как ДНК-полимера- зы, так и РНК-полимеразы самого разного происхождения: вирусов, бактерий, животных клеток. Механизм действия этих рифамицинов пока неясен. Известно, однако, что они, по-видимому, сильнее подавляют инициацию синтеза полинуклеотидных цепей, чем их элонгацию.
Источник: Ланчини Д., Паренти Ф., «Антибиотики. Пер. с англ. — М.: Мир. — 272 с., ил.» 1985