НОЦИЦЕПТИВНЫЕ СИСТЕМЫ
Ноцицептивный код, распространяясь по нейронным сетям мозга, претерпевает большие качественные и количественные изменения. Выделяются четыре качественно различных физиологических процесса, которые создает в мозге ноцицептивный код: 1) трансдукция; 2) трансмиссия; 3) модуляция;
Первичные афферентные чувствительные волокна, обеспечивающие генерацию ноцицептивного кода, называются ноцицепторами. Поскольку не удается структурно выделить ноцицепторы, найти какие-то специфические признаки, этот термин условно применяют для обозначения волокон и предполагаемых рецепторов, ответственных за формирование боли. Категории ноцицепторов представлены в таблице 4.
Ноцицептивное кодирование связывают с волокнами класса А8 и С (табл. 3). Большинство волокон типа А8 и С завершаются не специальными структурами, а как свободные нервные окончания. При иннервации кожи, волокна типа А?» теряют свою миелиновую оболочку, их аксон остается окружен-
Категории ноцицепторов
ным собственной базальной мембраной и швановскими клетками вплоть до их окончания в эпидермисе.
Волокна типа С могут оканчиваться в поверхностных слоях кожи в виде свободных кистеобразных формирований, окруженных цитоплазмой швановских клеток (в волосистой части кожи) или как вертикально ориентированные волокна, образуя окончания типа точек в поверхностных слоях дермы (безволосая гладкая кожа) — табл. 5
Классификация и характеристики волокон периферических нервов
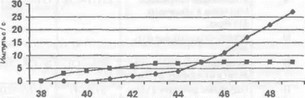
Проведенные специальные исследования позволяют сделать вывод, что свободные нервные окончания формируют ноцицептивный код. Важной функциональной особенностью ноцицепторов, является изменение частоты ответной реакции пропорционально силе действующего стимула. Изменение частоты при температурном воздействии показано на рис. 12.
Генерация ноцицептивного кода А8-волокнами происходит следующим образом. При нарастающей интенсивности раздражения, возбудимость А8-волокон резко возрастает, это отличает ноцицептивную афферентацию от неноцицептивной, т.е. имеется более высокий порог стимуляции. Поэтому волокна А8 называют механорецепторами высокого порога. Эти рецепторы не реагируют на однократную термальную стимуляцию, однако после повторных стимуляций они приобретают повышенную чувствительность и усиливают частоту своих сигналов. Подобный процесс известен как сенсибилизация. Примерно 20—50% А8-ноцицепторов в нормальных условиях реагируют как на термальные, так и на механические стимулы без сенсибилизации, но поддаются и сенсибилизации.
В отличие от А8-волокон С волокна генерируют ноцицептивный код при раздражении механического, термического и химического характера. Их еще называют С-полимодальны- ми ноцицепторами. Их рецептивные поля могут быть достаточно большими (до 17 мм2). Этим они отличаются от А5-но- цицепторов, рецептивные поля которых обычно представля-
Температура, С
#— —терминальные ноцицептор! ; —а неноцицептивные терморецепторы
Рис. 12. Сопоставление реакции различных рецепторов
ют собой группу точек. С-полимодальные ноцицепторы приобретают чувствительность после повторных раздражений и могут выдавать продолжительные сигналы. Так, химические раздражители продуцируют сигналы, длящиеся несколько минут.
Исследованиями установлено, что однократное раздражение Аб-волокон вызывает ощущение покалывания, а повторное раздражение — боль Воздействие на С-волокна вызывает чувство интенсивной и длительной бопи.
Волокна разных типов блокируются различными воздействиями (дгвление, местные анестетики и т.д.) по-разному. Местные анестетики в очень низкой концентрации блокируют, преимущественно, очень малые немиелинизированные волокна, в то время как при надавливании прежде всего блокируются миелинизированные волокна. Восприятие боли сохраняется на фоне примененного давления. Эта оставшаяся часть восприятия боли связана с активацией С-волокон.
При повреждении кожи важны процессы сенсибилизации и гипералгезии. В начале века Lewis описал характерную триаду после повреждения кожи: 1) интенсивное расширение сосудов; 2) местный отек; 3) последующее расширение сосудов в окружности места раздражения (воспаление). При этом больной отмечает снижение порога чувствительности к неповреждающим воздействиям (гиперестезия), снижение порога к повреждающим воздействиям и усиление боли в ответ на повреждающее раздражение (первичная гипералгезия), нанесенное в зоне поражения. Эти нарушения быстро распространяются на соседние участки, не подвергшиеся воздействию (вторичная гипералгезия). Вторичная гипералгезия зависит от активности первичных немиелинизированных афферентов при сенсибилизации С-полимодальных ноцицепторов (С-ПМН). Сама сенсибилизация С-ПМН и спонтанная деполимеризация развиваются после повреждающего раздражения даже вне их рецептивных полей. Эта активность С-ПМН в неповрежденных тканях служит причиной распространенной вазодилатации, отека и дальнейшей сенсибилизации С-ПМН в соседних рецептивных полях. Подобное явление получило название «не
врогенное воспаление» из-за его сходства с воспалительным процессом.
Гиперачгезию, относящуюся к реакции больного, не следует смешивать с сенсибилизацией, которая является реакцией ноцицептивных афферентных волокон.
В зоне поражения в процессе воздействия накапливаются ноциогенные вещества (табл 6). Источники происхождения этих веществ различны, в том числе поврежденные клетки, ноцицепторы, повышенная проницаемость капилляров и повышенная продукциях этих веществ энзимами.
Процесс трансдукции может инициироваться и усиливаться в результате действия нескольких механизмов: 1) прямая активация ноцицепторов; 2) сенсибилизация ноцицепторов с последующим усилением их активности; 3) выход ноциоген- ных веществ из плазмы. Процесс трансдукции заканчивается генерацией различных вариантов ноцицептивного кода. Харак-
Таблица 6
Алгогенные субстанции, вовлеченные в трансдукцию
тер кода зависит от вида повреждающего воздействия. Далее ноцицептивный код проводится в вышележащие отделы нервной системы.
В результате исследований, начатых в 50-х годах Рексе- дом, выяснено, что спинной мозг по своей ультраструктуре разделяется на ряд слоев, называемых пластинами (рис. 13) В пластинах, называемых еще пластинами Рекседа, находятся как возбуждающие, так и тормозящие промежуточные нейроны, а также передающие клетки, транслирующие ноцицептивный код в ростральные отделы мозга. Часть нейронов в этих пластинах реагирует только на ноциогенные стимулы, другие восприимчивы к широкому спектру стимулов Волокна типа Аб и С входят в спинной мозг в вентральном боковом участке
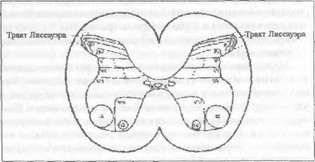
Рис. 13. Пластины Рекседа
заднего корешка, вступая затем в состав тракта Лиссауэра и заканчиваясь синапсами в пластинах I, II и V.
Основным путем передачи болевой информации в направлении коры головного мозга является спинно-таламический путь. В свою очередь, он подразделяется на неоспинно-тала- мический и палеоспинно-таламический (Albe-Fessard D., Fessard А., 1975, Belanger A.Y., 1985). Тела клеток спинно-таламического пути расположены преимущественно в пластинах I и V, а также VII и VIII. Восходящие аксоны этих нейронов проходят в передне-боковых канатиках спинного мозга. На уровне таламуса из этого квадранта выделяются два разных типа проводящих путей. Латеральная их часть (неоспин- но-таламический путь) образует синапсы в латеральном отделе таламуса, откуда сигналы проецируются в соматосенсорную зону коры. Латеральный спинно-таламический путь обеспечивает сенсорно-дискриминантные аспекты болевой перцепции. Неоспинно-таламический путь, филогенетически более молодой, состоит из длинных волокон, расположенных в наружных отделах спинно-таламического пути. Четкая сомато- топическая организация позволяет ему осуществлять передачу болевой и тактильной информации о положении периферического стимула в пространстве и времени, его интенсивности и продолжительности, т.е. дискриминационные аспекты боли. Через него в основном осуществляется передача острой (хорошо локализованной) боли Неоспинно-таламический путь заканчивается в ядрах вентролатерального и заднего таламуса, тесно связанных с первой и второй сомато-сенсорны- ми зонами коры.
Медиальный спинно-таламический (палеоспинно-талами- ческий) путь образует многочисленные синапсы в ретикулярной формации ствола мозга, в медиальном отделе таламуса, в периаквед) ктальном сером веществе и в гипоталамусе. Последующая проекция происходит диффузно в разные зоны коры и в лимбическую систему, принимает участие в передаче информации и формировании ощущения тупой, слабо локализованной хронической боли, т.е. недискриминантных ее аспектов. В связи с широким распространением окончаний в меди
альных и интраламинарных ядрах таламуса, а также гипоталамуса, структурах лимбического комплекса спинно-таламический путь приспособлен для формирования разнообразных супрасегментарных рефлекторных ответов, включающих изменение дыхания, кровообращения, эндокринных функций, мотивационно-поведенческие и сложные защитные функции. Кроме того, полагают, что импульсная активность в палеоспин- но-таламическом пути активирует нисходящую систему подавления боли, связанную с эндогенными агентами (Belanger A.Y., 1985). Таким образом, медиальный спинно-таламический путь обеспечивает аффективно-мотивационные аспекты болевой перцепции.
Следовательно, спинно-таламический путь через вентро- базальные ядра тачамуса образует специфическую систему боли, а через интраламинарные и медиальные ядра — неспецифическую. Разрушение первых образований проявляется в клинике аналгезией и нарушением дискриминационной чувствительности, разрушение вторых — крайне мучительными с оттенком жжения «таламическими» болями в контралатеральной половине туловища, лица, дистальных отделов конечностей. Считается, что таламус является конечной «станцией» переключения болевой импульсации (Comelli F., 1975).
Волокна третьего нейрона всех видов чувствительности идут от таламуса через заднюю ножку внутренней капсулы к задней центральной извилине и к верхней теменной дольке. По данным В.К.Решетняка и соавт. (1986), первая соматосенсорная зона связана с тонким дискриминационным анализом соматической чувствительности и участвует в восприятии боли. Вторая сенсомоторная зона отвечает за первичный ситуационный анализ и связана с регуляцией двигательного ответа на боль. Примечательно, что разрушение первой зоны приводит к повышению порога болевой чувствительности на противоположной стороне, а разрушение второй, напротив, снижает порог болевой чувствительности. Болевая афферентация может поступить в головной мозг непосредственно через чувствительные клетки основания заднего рога, которые соединяются между собой вставочными нейронами (клетки-передатчики)
Известен также «окольный» путь передачи боли вначале через пограничную симпатическую цепочку, а затем в верхних ее отделах болевое возбуждение вступает в спинной мозг через задние корешки. По данным R.Melzak и P.Wall (1965), одним из модуляторов болевого раздражения могут быть задние столбы спинного мозга. Кроме этого установлено, что на всех релейных участках передачи болевых импульсов (в первичных болевых афферентах, задних рогах спинного мозга, таламусе, лимбико-ретикулярном комплексе, в небольших количествах в коре головного мозга) обнаружено наличие нейропептида — субстанции Р — (pain — боль). Субстанция Р(состоит из 11 аминокислот) и может быть передатчиком болевых стимулов. Вещество Р выделяется только при интенсивном болевом воздействии. Субстанция Р может взаимодействовать и с катехоламинами, она обладает способностью защищать организм от стрессов, может вызывать снижение АД (Oehme Р. е.а., 1986).
Таким образом, процесс трансмиссии связан с вов лечением в ноцицепцию самых различных отделов нервной системы. Однако в узловых местах нейронная сеть меняет характеристики ноцицептивного кода и соответственно сам процесс формирования феномена боли.
Издавна обращалось внимание на то, что повреждения одинаковой тяжести вызывают у разных лиц резко различающиеся по степени выраженности болевые ощущения в зависимости от физиологических, ситуационных, этнических, культурных и других факторов. Травма на поле боя может сопровождаться значительно меньшими болями по сравнению с подобными же повреждениями в обычных условиях. У ряда больных введение физиологического раствора может обеспечить выраженное обезболивание. Все это говорит о том, что ЦНС способна модулировать боль, и механизмы этого процесса в последние годы начали выясняться.
Reynolds (1969) первым описал глубокую аналгезию у крыс во время электрической стимуляции центрального серого вещества, окружающего водопровод мозга.
Другим механизмом модуляции является наличие эндогенных опиоидов. Многие зоны мозга у человека и животных способны при стимуляции вызывать аналгезию. Обнаружено, что такие участки совпадают или перекрывают зоны мозга с повышенной концентрацией эндогенных опиоидных нейротрансмиттеров. Многочисленные исследования не выявили каких-либо отличий в структуре, физиологии и фармакологии при аналгезии, вызванной стимуляцией или назначением опиоидов. Введение минимальн ых количеств морфина в зоны мозга, раздражение которых продуцировало аналгезию, вызывало выраженное обезболивание.
Важными центрами нисходящей модулирующей системы являются перивентрикулярное и периакведуктальное скопление серого вещества, дорсолатеральный отдел моста, ядро шва и ростровентральный отдел продолговатого мозга. Биогенные амины (серотонин и норадреналин), так же как и эндогенные опиоиды, представляют собой нейротрансмиттеры этой системы. Антиноцицептивные системы оказывают существенное влияние на модуляцию.
Указанные выше процессы имеют механический харакеер и не отличаются от процессов, лежащих в основе любого другого вида восприятия. Они составляют один из компонентов сенсорно-дискриминативных аспектов боли.
Важной проблемой, затрудняющей понимание боли, яв
ляется отсутствие развитого структурно-логического языка феномена боли. В клинической медицине и неврологии боль рассматривается как единый феномен, качественные стороны которого анализируются Развитие структурно-логического «языка» боли требуют новых понятий, и введение ноцицептивной системы является началом этого процесса. Следующими необходимо определить понятия: ноциогенная зона, ноци- огенная структура. Ноциогенная зона — это спонтанно возникающая и исчезающая зона, которая продуцирует ноциоген- ный код. Ноциогенные структуры — это различные структуры организма, которые приобретают свойство постоянно или периодически продуцировать ноциогенный код. Ноциогенные структуры могут объединяться в ноциогенные зоны. Например, при тригеминальном болевом синдроме ноциогенными структурами системы тройничного нерва будут, корешок, нервные волокна, звездчатый узел, ядра ствола мозга, вегетативные ганглии лица. Важно отметить, что ноциогенная структура — это обобщенное понятие, она может, условно говоря, «мигрировать», т.е. это — свойство, а свойство может появляться у различных тканей. Отсюда можно объяснить хронизацию боли, которая связана с тем, что постепенно ноциогенными структурами становятся структуры нервной системы. И если сначала ноциогенной структурой является соединительная ткань, то затем свойство ноциогенности приобретают структуры нервной системы — нервные волокна в корешке, затем нервные клетки задних рогов спинного мозга. Согласно экспериментальным данным Г.Н.Крыжановского, формируются генераторы патологически усиленной импульсации, которые и являются ноциогенными структурами в нервной системе. Различные методы лечения направлены на устранение ноциогенных структур (терапевтические методы приводят к трансформации, т е. к устранению свойства ноциогенности у тканей, хирурги ческие методы направлены на устранение ноциогенных структур)-
Очень часто ноциогенной структурой является мышца. Мышечная ткань не обладает болевой чувствительностью, т.е. бочевое ощущение не возникает при уколе, разрезе мышцы и
других механических воздействиях. Но мышечная боль существует и испытывается довольно часто. Моделирование мышцы позволяет понять, при каких условиях мышца становится ноциогенной структурой. В соответствии с двойным реципрок- ным принципом иннервации и регуляции чрезмерное усиление тонуса мышечных волокон, т.е. их спазмирование, при котором возникают болевые ощущения, возможно только при нарушении баланса между холинергической и адренергической составляющими в сторону усиления первой. В этом случае при сильном сокращении мышечных волокон происходит сильное растяжение тензорецепторов. В результате с них возникает афферентный сигнал, который оказывает стимулирующее воздействие на ретикулярную формацию, которая и формирует ноцицептивный код и затем болевое ощущение. И наоборот, при чрезмерном ослаблении тонуса мышечных волокон по той или иной причине они растягиваются, а с мышечных веретен возникает афферентный сигнал большой силы, который опять же через ретикулярную формацию генерирует болевое ощущение.
В клиническом аспекте, боль мобилизует самые разнообразные функциональные системы для защиты организма от воздействия вредного фактора, включая такие компоненты, как эмоции, соматические, поведенческие и вегетативные реакции, мотивации, память.
Ноцицептивный код прогрессивно вовлекает в процесс возбуждения различные структуры мозга, что становится особенно заметно на ассоциативных связях с концептуальной полифакторной моделью боли, рассматривающей ее как последовательность генерализации негативно очерченного процесса с нарастающим числом составляющих:
Современные представления о функционировании ноци- цептивной системы имеют не только академический интерес. Важное практическое значение эти знания приобретают в формировании рациональных подходов к устранению хронических болевых синдромов. Процессы трансдукции, трансмиссии и модуляции протекают одновременно, влияя на ноцицептив- ные нейроны в пластинах Рекседа (I, II, V). Ноцицепция является динамичным и изменчивым процессом, подверженным модификации и модуляции на разных уровнях с участием высокоспецифических факторов. Варианты устранения боли могут быть различными (рис. 14) и зависят от структурно-информационного строения болевого синдрома.
- перцепция.
- Трансдукция — это динамический процесс, при котором воздействие, выходящее за пределы физиологических границ, трансформируется в виде электрической активности на окончаниях чувствительных нервов, формируя информационный ноцицептивный код.
Первичные афферентные чувствительные волокна, обеспечивающие генерацию ноцицептивного кода, называются ноцицепторами. Поскольку не удается структурно выделить ноцицепторы, найти какие-то специфические признаки, этот термин условно применяют для обозначения волокон и предполагаемых рецепторов, ответственных за формирование боли. Категории ноцицепторов представлены в таблице 4.
Ноцицептивное кодирование связывают с волокнами класса А8 и С (табл. 3). Большинство волокон типа А8 и С завершаются не специальными структурами, а как свободные нервные окончания. При иннервации кожи, волокна типа А?» теряют свою миелиновую оболочку, их аксон остается окружен-
Категории ноцицепторов
|
Миелинизация |
Классификация Гассера и Эрлангера |
Порог чувстви- течьности |
Сенсиби лизация |
Рецептивные поля |
|
Миелинизиро- ванные механорецепторы высокого порога |
А5 |
Высокий (повреж дающий) интенсив ный |
Да |
Малые |
|
Механотер- мальные |
А5 |
То же |
|
(( |
|
Немиелинизи- рованные С- полимодаль- ные ноцицеп- торы Неноцицепто- ры, миелини- зированные |
С |
lt;« |
|
Большие |
|
Механорецептор низкого порога (не относится к но- цицепторам, но приводится для сравнения) |
Аа |
Низкий |
Нет |
|
ным собственной базальной мембраной и швановскими клетками вплоть до их окончания в эпидермисе.
Волокна типа С могут оканчиваться в поверхностных слоях кожи в виде свободных кистеобразных формирований, окруженных цитоплазмой швановских клеток (в волосистой части кожи) или как вертикально ориентированные волокна, образуя окончания типа точек в поверхностных слоях дермы (безволосая гладкая кожа) — табл. 5
Классификация и характеристики волокон периферических нервов
|
Волокна группы |
Иннервация |
Средний диаметр, мкм |
Средняя скорость проведения, м/с |
|
Классификация Гассера и Эрпангера (афференты эфференты) |
|||
|
Аа |
Первичные моторные к скелетным мышцам (миелини- зированные) |
15(12—20) |
100(70— 120) |
|
АР |
Кожные афференты, тактильные v прессорное восприятие (миелинизирован- ные) |
8 (5—15) |
50 (30—70) |
|
Ау |
Моторные к мышцам (мие- линизированные) |
6(6—8) |
20 (15—30) |
|
А5 |
Механорецепторы, ноцицеп- торы (миелинизированные) |
lt;3(1—4) |
15 (12—30) |
|
В |
Симпатические преганглионарные (миелинизированные) |
3 (1—3) |
7 (3—15) |
|
С |
Механорецепторы, ноцицеп- торы, симпатические пост- ганглионарные (немиелини- зированные) |
1(0,5-1,5) |
1 (0,5-2) |
|
Классификация Ллойда - Ханта (только мышечные афАеренты) |
|||
|
1а |
Кольцевидно-спиральные окончания в мышечных футлярах |
13(11—20) |
75(70—120) |
|
1Ъ |
Нервно-сухожильные футляры |
|
|
|
II |
Окончания как «цветочная роса» в мышечных футлярах |
9(4—12) |
55 (25—70) |
|
III |
Ощущение давления в ноци- цепторах мышц |
3(1-4) |
11 (10—25) |
|
IV |
Немиелинизированные волокна С, механические но- цицепторы |
1 (0,5—1,5) |
1 (0,5—2) |
Проведенные специальные исследования позволяют сделать вывод, что свободные нервные окончания формируют ноцицептивный код. Важной функциональной особенностью ноцицепторов, является изменение частоты ответной реакции пропорционально силе действующего стимула. Изменение частоты при температурном воздействии показано на рис. 12.
Генерация ноцицептивного кода А8-волокнами происходит следующим образом. При нарастающей интенсивности раздражения, возбудимость А8-волокон резко возрастает, это отличает ноцицептивную афферентацию от неноцицептивной, т.е. имеется более высокий порог стимуляции. Поэтому волокна А8 называют механорецепторами высокого порога. Эти рецепторы не реагируют на однократную термальную стимуляцию, однако после повторных стимуляций они приобретают повышенную чувствительность и усиливают частоту своих сигналов. Подобный процесс известен как сенсибилизация. Примерно 20—50% А8-ноцицепторов в нормальных условиях реагируют как на термальные, так и на механические стимулы без сенсибилизации, но поддаются и сенсибилизации.
В отличие от А8-волокон С волокна генерируют ноцицептивный код при раздражении механического, термического и химического характера. Их еще называют С-полимодальны- ми ноцицепторами. Их рецептивные поля могут быть достаточно большими (до 17 мм2). Этим они отличаются от А5-но- цицепторов, рецептивные поля которых обычно представля-
Температура, С
#— —терминальные ноцицептор! ; —а неноцицептивные терморецепторы
Рис. 12. Сопоставление реакции различных рецепторов
ют собой группу точек. С-полимодальные ноцицепторы приобретают чувствительность после повторных раздражений и могут выдавать продолжительные сигналы. Так, химические раздражители продуцируют сигналы, длящиеся несколько минут.
Исследованиями установлено, что однократное раздражение Аб-волокон вызывает ощущение покалывания, а повторное раздражение — боль Воздействие на С-волокна вызывает чувство интенсивной и длительной бопи.
Волокна разных типов блокируются различными воздействиями (дгвление, местные анестетики и т.д.) по-разному. Местные анестетики в очень низкой концентрации блокируют, преимущественно, очень малые немиелинизированные волокна, в то время как при надавливании прежде всего блокируются миелинизированные волокна. Восприятие боли сохраняется на фоне примененного давления. Эта оставшаяся часть восприятия боли связана с активацией С-волокон.
При повреждении кожи важны процессы сенсибилизации и гипералгезии. В начале века Lewis описал характерную триаду после повреждения кожи: 1) интенсивное расширение сосудов; 2) местный отек; 3) последующее расширение сосудов в окружности места раздражения (воспаление). При этом больной отмечает снижение порога чувствительности к неповреждающим воздействиям (гиперестезия), снижение порога к повреждающим воздействиям и усиление боли в ответ на повреждающее раздражение (первичная гипералгезия), нанесенное в зоне поражения. Эти нарушения быстро распространяются на соседние участки, не подвергшиеся воздействию (вторичная гипералгезия). Вторичная гипералгезия зависит от активности первичных немиелинизированных афферентов при сенсибилизации С-полимодальных ноцицепторов (С-ПМН). Сама сенсибилизация С-ПМН и спонтанная деполимеризация развиваются после повреждающего раздражения даже вне их рецептивных полей. Эта активность С-ПМН в неповрежденных тканях служит причиной распространенной вазодилатации, отека и дальнейшей сенсибилизации С-ПМН в соседних рецептивных полях. Подобное явление получило название «не
врогенное воспаление» из-за его сходства с воспалительным процессом.
Гиперачгезию, относящуюся к реакции больного, не следует смешивать с сенсибилизацией, которая является реакцией ноцицептивных афферентных волокон.
В зоне поражения в процессе воздействия накапливаются ноциогенные вещества (табл 6). Источники происхождения этих веществ различны, в том числе поврежденные клетки, ноцицепторы, повышенная проницаемость капилляров и повышенная продукциях этих веществ энзимами.
Процесс трансдукции может инициироваться и усиливаться в результате действия нескольких механизмов: 1) прямая активация ноцицепторов; 2) сенсибилизация ноцицепторов с последующим усилением их активности; 3) выход ноциоген- ных веществ из плазмы. Процесс трансдукции заканчивается генерацией различных вариантов ноцицептивного кода. Харак-
Таблица 6
Алгогенные субстанции, вовлеченные в трансдукцию
|
Субстанция |
Источник |
Фермент |
Влияние на первичный афферент |
|
Калий |
Поврежденные клетки |
— |
Активирование |
|
Серотонин |
Тромбоциты |
— |
(1 |
|
Брадикинин |
К”ниноген плазмы |
Калликоечн |
w |
|
Гистамин |
Тучные клетки |
— |
|
|
Простагланди- ны |
Арахидоновая кислота из поврежденных клеток |
Циклоокси- геназа |
Сенсибилизация |
|
Лейкотриены |
То же |
Липоксигеназа |
|
|
Субстанция Р |
Первичные афференты |
— |
(С |
тер кода зависит от вида повреждающего воздействия. Далее ноцицептивный код проводится в вышележащие отделы нервной системы.
- Трансмиссия — распространение ноцицептивного кода по системе чувствительных нервов. Невральные проводящие пути, обеспечивающие трансмиссию, сформированы из трех компонентов: 1) первичный чувствительный афферентный нейрон, доходящий до спинного мозга; 2) восходящий промежуточный нейрон, простирающийся от спинного мозга до стволовой части головного мозга и таламуса; 3) таламокорти- кальные проекции.
В результате исследований, начатых в 50-х годах Рексе- дом, выяснено, что спинной мозг по своей ультраструктуре разделяется на ряд слоев, называемых пластинами (рис. 13) В пластинах, называемых еще пластинами Рекседа, находятся как возбуждающие, так и тормозящие промежуточные нейроны, а также передающие клетки, транслирующие ноцицептивный код в ростральные отделы мозга. Часть нейронов в этих пластинах реагирует только на ноциогенные стимулы, другие восприимчивы к широкому спектру стимулов Волокна типа Аб и С входят в спинной мозг в вентральном боковом участке
Рис. 13. Пластины Рекседа
заднего корешка, вступая затем в состав тракта Лиссауэра и заканчиваясь синапсами в пластинах I, II и V.
Основным путем передачи болевой информации в направлении коры головного мозга является спинно-таламический путь. В свою очередь, он подразделяется на неоспинно-тала- мический и палеоспинно-таламический (Albe-Fessard D., Fessard А., 1975, Belanger A.Y., 1985). Тела клеток спинно-таламического пути расположены преимущественно в пластинах I и V, а также VII и VIII. Восходящие аксоны этих нейронов проходят в передне-боковых канатиках спинного мозга. На уровне таламуса из этого квадранта выделяются два разных типа проводящих путей. Латеральная их часть (неоспин- но-таламический путь) образует синапсы в латеральном отделе таламуса, откуда сигналы проецируются в соматосенсорную зону коры. Латеральный спинно-таламический путь обеспечивает сенсорно-дискриминантные аспекты болевой перцепции. Неоспинно-таламический путь, филогенетически более молодой, состоит из длинных волокон, расположенных в наружных отделах спинно-таламического пути. Четкая сомато- топическая организация позволяет ему осуществлять передачу болевой и тактильной информации о положении периферического стимула в пространстве и времени, его интенсивности и продолжительности, т.е. дискриминационные аспекты боли. Через него в основном осуществляется передача острой (хорошо локализованной) боли Неоспинно-таламический путь заканчивается в ядрах вентролатерального и заднего таламуса, тесно связанных с первой и второй сомато-сенсорны- ми зонами коры.
Медиальный спинно-таламический (палеоспинно-талами- ческий) путь образует многочисленные синапсы в ретикулярной формации ствола мозга, в медиальном отделе таламуса, в периаквед) ктальном сером веществе и в гипоталамусе. Последующая проекция происходит диффузно в разные зоны коры и в лимбическую систему, принимает участие в передаче информации и формировании ощущения тупой, слабо локализованной хронической боли, т.е. недискриминантных ее аспектов. В связи с широким распространением окончаний в меди
альных и интраламинарных ядрах таламуса, а также гипоталамуса, структурах лимбического комплекса спинно-таламический путь приспособлен для формирования разнообразных супрасегментарных рефлекторных ответов, включающих изменение дыхания, кровообращения, эндокринных функций, мотивационно-поведенческие и сложные защитные функции. Кроме того, полагают, что импульсная активность в палеоспин- но-таламическом пути активирует нисходящую систему подавления боли, связанную с эндогенными агентами (Belanger A.Y., 1985). Таким образом, медиальный спинно-таламический путь обеспечивает аффективно-мотивационные аспекты болевой перцепции.
Следовательно, спинно-таламический путь через вентро- базальные ядра тачамуса образует специфическую систему боли, а через интраламинарные и медиальные ядра — неспецифическую. Разрушение первых образований проявляется в клинике аналгезией и нарушением дискриминационной чувствительности, разрушение вторых — крайне мучительными с оттенком жжения «таламическими» болями в контралатеральной половине туловища, лица, дистальных отделов конечностей. Считается, что таламус является конечной «станцией» переключения болевой импульсации (Comelli F., 1975).
Волокна третьего нейрона всех видов чувствительности идут от таламуса через заднюю ножку внутренней капсулы к задней центральной извилине и к верхней теменной дольке. По данным В.К.Решетняка и соавт. (1986), первая соматосенсорная зона связана с тонким дискриминационным анализом соматической чувствительности и участвует в восприятии боли. Вторая сенсомоторная зона отвечает за первичный ситуационный анализ и связана с регуляцией двигательного ответа на боль. Примечательно, что разрушение первой зоны приводит к повышению порога болевой чувствительности на противоположной стороне, а разрушение второй, напротив, снижает порог болевой чувствительности. Болевая афферентация может поступить в головной мозг непосредственно через чувствительные клетки основания заднего рога, которые соединяются между собой вставочными нейронами (клетки-передатчики)
Известен также «окольный» путь передачи боли вначале через пограничную симпатическую цепочку, а затем в верхних ее отделах болевое возбуждение вступает в спинной мозг через задние корешки. По данным R.Melzak и P.Wall (1965), одним из модуляторов болевого раздражения могут быть задние столбы спинного мозга. Кроме этого установлено, что на всех релейных участках передачи болевых импульсов (в первичных болевых афферентах, задних рогах спинного мозга, таламусе, лимбико-ретикулярном комплексе, в небольших количествах в коре головного мозга) обнаружено наличие нейропептида — субстанции Р — (pain — боль). Субстанция Р(состоит из 11 аминокислот) и может быть передатчиком болевых стимулов. Вещество Р выделяется только при интенсивном болевом воздействии. Субстанция Р может взаимодействовать и с катехоламинами, она обладает способностью защищать организм от стрессов, может вызывать снижение АД (Oehme Р. е.а., 1986).
Таким образом, процесс трансмиссии связан с вов лечением в ноцицепцию самых различных отделов нервной системы. Однако в узловых местах нейронная сеть меняет характеристики ноцицептивного кода и соответственно сам процесс формирования феномена боли.
- Модуляция - это процесс, при котором ноцицеп- тивная информация модифицируется при трансмиссии по но- цицептивной системе
Издавна обращалось внимание на то, что повреждения одинаковой тяжести вызывают у разных лиц резко различающиеся по степени выраженности болевые ощущения в зависимости от физиологических, ситуационных, этнических, культурных и других факторов. Травма на поле боя может сопровождаться значительно меньшими болями по сравнению с подобными же повреждениями в обычных условиях. У ряда больных введение физиологического раствора может обеспечить выраженное обезболивание. Все это говорит о том, что ЦНС способна модулировать боль, и механизмы этого процесса в последние годы начали выясняться.
Reynolds (1969) первым описал глубокую аналгезию у крыс во время электрической стимуляции центрального серого вещества, окружающего водопровод мозга.
Другим механизмом модуляции является наличие эндогенных опиоидов. Многие зоны мозга у человека и животных способны при стимуляции вызывать аналгезию. Обнаружено, что такие участки совпадают или перекрывают зоны мозга с повышенной концентрацией эндогенных опиоидных нейротрансмиттеров. Многочисленные исследования не выявили каких-либо отличий в структуре, физиологии и фармакологии при аналгезии, вызванной стимуляцией или назначением опиоидов. Введение минимальн ых количеств морфина в зоны мозга, раздражение которых продуцировало аналгезию, вызывало выраженное обезболивание.
Важными центрами нисходящей модулирующей системы являются перивентрикулярное и периакведуктальное скопление серого вещества, дорсолатеральный отдел моста, ядро шва и ростровентральный отдел продолговатого мозга. Биогенные амины (серотонин и норадреналин), так же как и эндогенные опиоиды, представляют собой нейротрансмиттеры этой системы. Антиноцицептивные системы оказывают существенное влияние на модуляцию.
Указанные выше процессы имеют механический харакеер и не отличаются от процессов, лежащих в основе любого другого вида восприятия. Они составляют один из компонентов сенсорно-дискриминативных аспектов боли.
- Перцепция является финальным процессом, при котором трансдукция, трансмиссия и модуляция, взаимодействуя с индивидуальными физиологическими особенностями личности, создают конечное субъективное эмоциональное ощущение, воспринимаемое нами как боль. Боль — это определенное состояние организма, возникающее под действием болевых информационных (ноциогенных) кодов, формирующихся в периферической нервной системе в результате болевых стимулов или спонтанно в центральной нервной системе.
Важной проблемой, затрудняющей понимание боли, яв
ляется отсутствие развитого структурно-логического языка феномена боли. В клинической медицине и неврологии боль рассматривается как единый феномен, качественные стороны которого анализируются Развитие структурно-логического «языка» боли требуют новых понятий, и введение ноцицептивной системы является началом этого процесса. Следующими необходимо определить понятия: ноциогенная зона, ноци- огенная структура. Ноциогенная зона — это спонтанно возникающая и исчезающая зона, которая продуцирует ноциоген- ный код. Ноциогенные структуры — это различные структуры организма, которые приобретают свойство постоянно или периодически продуцировать ноциогенный код. Ноциогенные структуры могут объединяться в ноциогенные зоны. Например, при тригеминальном болевом синдроме ноциогенными структурами системы тройничного нерва будут, корешок, нервные волокна, звездчатый узел, ядра ствола мозга, вегетативные ганглии лица. Важно отметить, что ноциогенная структура — это обобщенное понятие, она может, условно говоря, «мигрировать», т.е. это — свойство, а свойство может появляться у различных тканей. Отсюда можно объяснить хронизацию боли, которая связана с тем, что постепенно ноциогенными структурами становятся структуры нервной системы. И если сначала ноциогенной структурой является соединительная ткань, то затем свойство ноциогенности приобретают структуры нервной системы — нервные волокна в корешке, затем нервные клетки задних рогов спинного мозга. Согласно экспериментальным данным Г.Н.Крыжановского, формируются генераторы патологически усиленной импульсации, которые и являются ноциогенными структурами в нервной системе. Различные методы лечения направлены на устранение ноциогенных структур (терапевтические методы приводят к трансформации, т е. к устранению свойства ноциогенности у тканей, хирурги ческие методы направлены на устранение ноциогенных структур)-
Очень часто ноциогенной структурой является мышца. Мышечная ткань не обладает болевой чувствительностью, т.е. бочевое ощущение не возникает при уколе, разрезе мышцы и
других механических воздействиях. Но мышечная боль существует и испытывается довольно часто. Моделирование мышцы позволяет понять, при каких условиях мышца становится ноциогенной структурой. В соответствии с двойным реципрок- ным принципом иннервации и регуляции чрезмерное усиление тонуса мышечных волокон, т.е. их спазмирование, при котором возникают болевые ощущения, возможно только при нарушении баланса между холинергической и адренергической составляющими в сторону усиления первой. В этом случае при сильном сокращении мышечных волокон происходит сильное растяжение тензорецепторов. В результате с них возникает афферентный сигнал, который оказывает стимулирующее воздействие на ретикулярную формацию, которая и формирует ноцицептивный код и затем болевое ощущение. И наоборот, при чрезмерном ослаблении тонуса мышечных волокон по той или иной причине они растягиваются, а с мышечных веретен возникает афферентный сигнал большой силы, который опять же через ретикулярную формацию генерирует болевое ощущение.
В клиническом аспекте, боль мобилизует самые разнообразные функциональные системы для защиты организма от воздействия вредного фактора, включая такие компоненты, как эмоции, соматические, поведенческие и вегетативные реакции, мотивации, память.
Ноцицептивный код прогрессивно вовлекает в процесс возбуждения различные структуры мозга, что становится особенно заметно на ассоциативных связях с концептуальной полифакторной моделью боли, рассматривающей ее как последовательность генерализации негативно очерченного процесса с нарастающим числом составляющих:
- ноцицепция (импульсация от рецептивного поля, передающаяся по Аб и С-афферентам);
- боль (интеграция ноцицептивных сигналов на уровне спинного мозга);
- страдание (негативное ощущение, генерированное в ЦНС и модулированное эмоциональными ситуациями, такими как страх, стресс, тревожность, депрессия);
- «болевое поведение» (моторно-мотивационный ответ организма, регулируемый всеми составляющими).
Современные представления о функционировании ноци- цептивной системы имеют не только академический интерес. Важное практическое значение эти знания приобретают в формировании рациональных подходов к устранению хронических болевых синдромов. Процессы трансдукции, трансмиссии и модуляции протекают одновременно, влияя на ноцицептив- ные нейроны в пластинах Рекседа (I, II, V). Ноцицепция является динамичным и изменчивым процессом, подверженным модификации и модуляции на разных уровнях с участием высокоспецифических факторов. Варианты устранения боли могут быть различными (рис. 14) и зависят от структурно-информационного строения болевого синдрома.