Теория саморегуляции эпидемического процесса
В. Д. Беляков с сотрудниками в 70-е гг. XX в. на основании собственных исследований и обобщения данных мировой науки сформулировал теорию (концепцию) внутренней саморегуляции эпидемического процесса, определяющей его саморазвитие. Главное в этой теории, что носит несомненно фундаментальный и универсальный характер, — это указание на то, что эпидемический процесс, т. е. межпопуляционное взаимоотношение паразита и хозяина, представляет собой систему, подчиняющуюся внутренним саморегуляционным процессам. Биология, медицина — в частности, уже давно установили, что каждый живой организм — это сложная саморегулирующаяся система, но за пределы организменного уровня представления о саморегуляции никогда не выходили.
Механизм, сущность саморегуляции, поданным авторов, слагается из следующих компонентов.
1. Популяции хозяина и паразита неоднородны по генотипным и фенотипным свойствам, характеризующим их отношение друг к другу.
Как известно, все единицы, любая составляющая материального мира не однородны, но авторы этот феномен определили как основную причину (базу) саморегуляторных процессов. В каждой популяции есть устойчивые и восприимчивые к инфекции люди, за счет последних обеспечивается существование паразита С другой стороны, в популяции паразита имеются вирулентные и невирулентные расы.
2. В эпидемическом процессе имеет место взаимообусловленная изменчивость биологических свойств обеих взаимодействующих популяций.
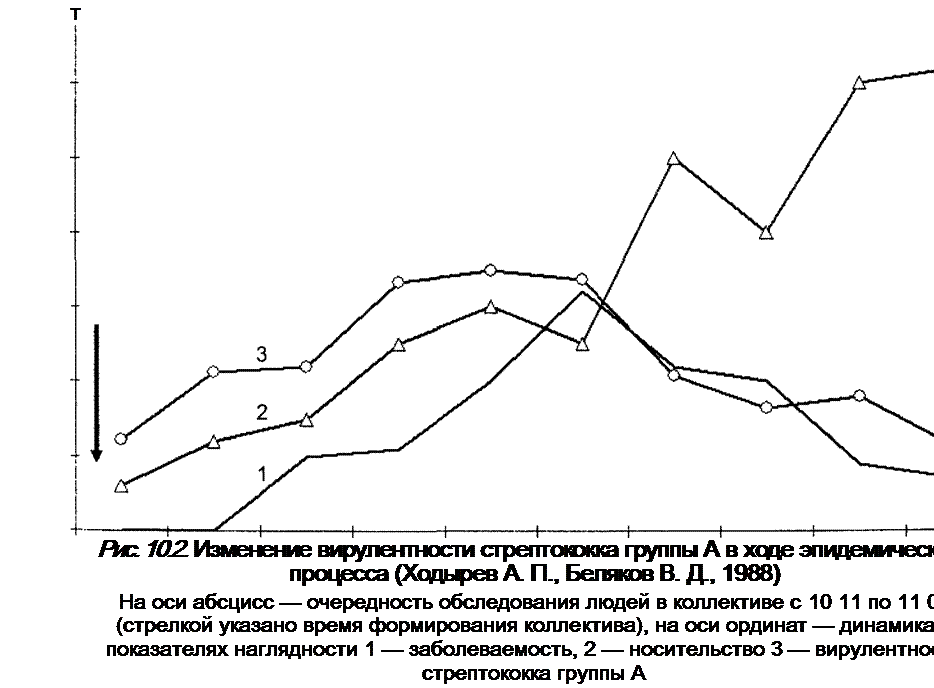
|
Выше приведены данные об изменениях среди населения при распространении заболеваний, передающихся воздушно-капельным путем: формирование популяционного иммунитета, оказывающего тормозящую роль в циркуляции возбудителя. Известны примеры изменчивости в ходе эпидемического процесса вирулентности возбудителя. Так, например, при дифтерии в эпидемический период в основном выделяются вирулентные разновидности возбудителя (тип гравис), в межэпидемический период — главным образом слабовирулентный тип митис. Авторы теории приводят убедительные материалы, характеризующие изменения в ходе развития эпидемического процесса вирулентности гемолитического стрептококка А (рис. 10 2). Из рисунка видно, что после формирования коллектива имел место подъем уровня носительства стрептококка группы А. Первые больные ангиной появились лишь через месяц. Параллельно отмечался рост вирулентности стрептококка группы А. Затем картина изменилась: синхронно снизилась заболеваемость ангиной и вирулентность стрептококка группы А, а носительство продолжало расти. Таким образом, авторы делают вывод о том, что изменение уровня заболеваемости в ходе эпидемического процесса связано с изменением вирулентности возбудителя.
На рис. 10.3 показано, как изменялась вирулентность синегнойной палочки, которая вызвала вспышку в урологическом стационаре
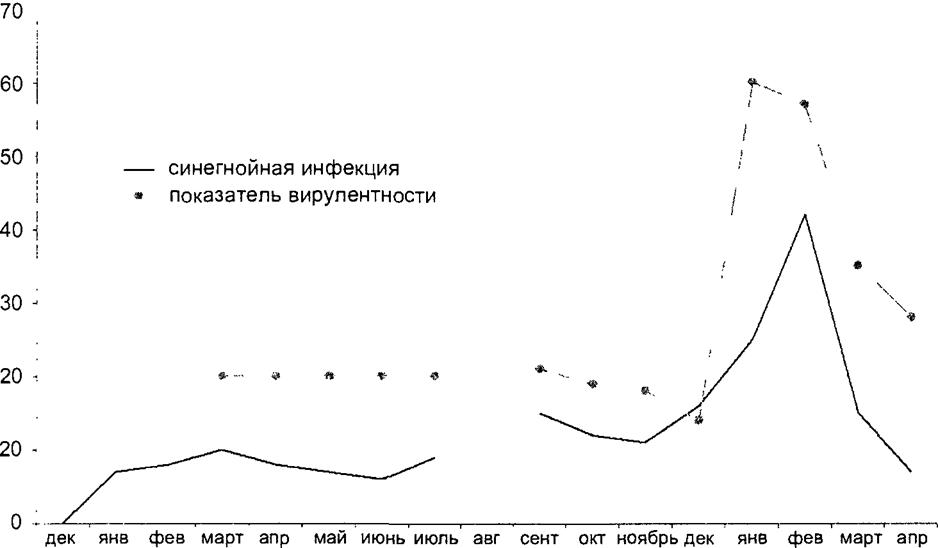
Рис. 10.3. Заболеваемость синегнойной инфекцией в урологическом стационаре и вирулентность выделенных штаммов (Л П. Зуева) |
На фоне регулярного выделения слабовирулентных рас синегнойной палочки перед вспышкой обнаружился подъем заболеваемости, который синхронизировался с ростом вирулентности выделяемых культур возбудителя, а снижение вирулентности выделенных культур сопровождалось постепенным снижением заболеваемости.
В пользу приведенного регуляторного механизма имеются данные при воздушно-капельных инфекциях в отношении популяции хозяина (формирование популяционного иммунитета), в отношении популяции паразита — при воздушно-капельных инфекциях, для которых характерно развитие инфекционного процесса как в манифестной, так и в бессимптомной форме, а также при заболеваниях, которые вызываются условно-патогенными бактериями. Однако универсальность этого механизма, участвующего в саморегуляции, требует дальнейшего изучения и подтверждения, поскольку имеются факты, не укладывающиеся в разбираемую схему Например, при кори (облигатно-патогенный паразит) заболевания, если не удается вмешаться в ход инфекционного процесса, как в эпидемический, так и в межэлидемический период протекают практически всегда тяжело (хотя, конечно, трудно представить, что вирус кори, обладающий неограниченными возможностями для быстрого размножения и смены поколений, не подвергался бы изменениям в ходе эпидемического процесса, по, возможно, изменяются иные свойства паразита) Не ясна возможность такого механизма регуляции при инфекциях пищеварительной системы, а также инфекциях, при которых сомнительна роль иммунитета (сифилис, гонорея, ВИЧ-инфекция, микозы, гель- ми нтозы и т д )
3. Имеет место фазовая самоперестройка популяции паразита, которая определяет неравномерность развития эпидемического процесса.
Именно это положение подвергалось резкой критике, хотя, не исключено, обсуждаемая теория в этой части, если отвлечься от постулируемой строгой последовательности числа и значимости фаз, в какой-то степени опередила время существующих представлений и стереотипов Авторы считают, что в популяции паразита имеется 4 последовательно сменяющих друг друга фазы* резервации, эпидемического преобразования, эпидемического распространения и, наконец, резервационного преобразования На примере острых воздушно-капельных инфекций авторами представлена принципиальная схема смены фаз (рис 10 4)
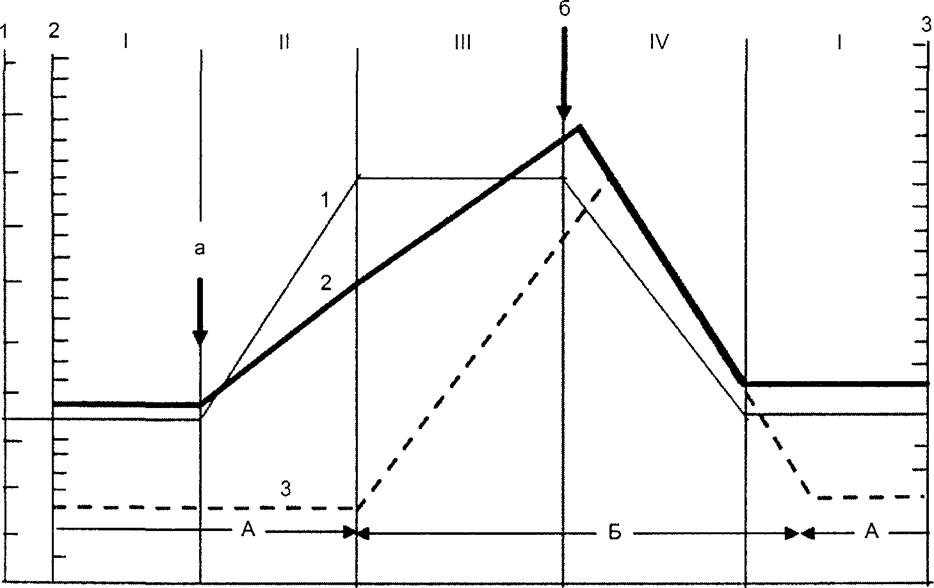
Рис. 10 4. Фазовые изменения популяции возбудителя (схема) (В. Д Беляков, Г Д Каминский, 1987 г ) На оси абсцисс — временные периоды, на оси ординат слева — показатели вирулентности (1) и численности (2) популяции возбудителя в условных единицах справа — показатели заболеваемости (3) в условных единицах Стрелками указано начало изменения среды обитания возбудителя а — достижение количественного порога восприимчивых б — достижение количественного порога иммунных в цепи циркуляции возбудителя Фазы I — резервация II — эпидемическое преобразование III — эпидемическое распространение IV — резервационное преобразование Периоды А — межсезонный (межэпидемический) Б — сезонный (эпидемический) |
Попытка распространить представленную схему на все инфекционные заболевания подверглась особенно сильной критике, и заставила авторов согласиться с возможностью отклонений от показанной на рис 104 и в табл ЮЛ картины Конечно, очень трудно во всех стучаях связать развитие эпидемического процесса с приведенными флювыми трансформациями паразита, их последовательностью и значимостью. Это относится к ряду кишечных инфекций, к инфекциях наружных покровов, многим хроническим инфекциям
Фазовые изменения популяции паразита
 (на примере инфекционно-иммунологических взаимоотношений с популяцией хозяина)
(на примере инфекционно-иммунологических взаимоотношений с популяцией хозяина)
Таблица 10.1
|
4. Влияние социальных и природных факторов на формирование и развитие эпидемического процесса, что общепризнано.
Саморегуляторные процессы в представленной В. Д. Беляковым с сотрудниками схеме очевидны при антропонозных воздушно-капельных инфекциях. При других группах антропонозов, нет сомнения, также действуют процессы саморегуляции, но их механизм иной (например, при брюшном тифе для обеспечения существования паразитического вида после перенесения острого инфекционного процесса формируется некоторая группа носителей, отдельные из них сохраняют возбудителя всю жизнь). Саморегуляторные процессы, в соответствии с собственными законами, сложившимися в эволюционном развитии, имеют место при зоонозных инфекциях (например, инкапсулирование личинок трихинелл в мышечной ткани хозяина).
В отношении факультативных паразитов также, по-видимому, можно говорить о возможности саморегуляции. При паразитировании среди людей саморегуляция ограничивается тем небольшим пространством, где обитают факультативные паразиты (например, отделения стационаров являются местом, Где складываются условия для сохранения за счет само- регуляционных процессов условно-патогенных микроорганизмов, причем это очевидно даже при активном воздействии на эти микроорганизмы). Надо отметить, что непаразитическая фаза (т. е. существование факультативного паразита во внешней среде) также находится в условиях саморегуляции, но факторы, которые обеспечивают сохранение агента, иные, адекватные внешней среде.
Итак, паразитарные системы существуют за счет саморегуляторных процессов, но характер их специфичен для каждой системы.
Источник: Л. П. Зуева, Р. X. Яфаев, «ЭПИДЕМИОЛОГИЯ» 2005
А так же в разделе «Теория саморегуляции эпидемического процесса »
- Глава 8 ОПРЕДЕЛЕНИЕ ЭПИДЕМИЧЕСКОГО ПРОЦЕСС
- 8.1. Характеристика паразитов
- Характеристика макроорганизма (хозяина)
- Глава 9 СОСТАВНЫЕ ЧАСТИ (ВНУТРЕННЯЯ СТРУКТУРА) ЭПИДЕМИЧЕСКОГО ПРОЦЕССА
- 9.1. Источник инфекции
- 9.2. Механизм передачи
- 9.2. Восприимчивый организм
- Глава 10 ПОПУЛЯЦИОННЫЙ ИММУНИТЕТ И ТЕОРИЯ САМОРЕГУЛЯЦИИ ЭПИДЕМИЧЕСКОГО ПРОЦЕССА
- Глава 11 КЛАССИФИКАЦИЯ ИНФЕКЦИОННЫХ БОЛЕЗНЕЙ
- Глава 19 ХАРАКТЕРИСТИКА ПРОЯВЛЕНИЙ ЭПИДЕМИЧЕСКОГО ПРОЦЕССА
- Глава 13 ФАКТОРЫ ЭПИДЕМИЧЕСКОГО ПРОЦЕССА
- Глава 14 ЭПИДЕМИОЛОГИЧЕСКИЙ НАДЗОР
- 14,1. Оперативный анализ
- 14.2. Ретроспективный анализ
- Глава 15 ПРОФИЛАКТИЧЕСКИЕ И ПРОТИВОЭПИДЕМИЧЕСКИЕ МЕРОПРИЯТИЯ
- 1.5.1. Общая структура профилактических и противоэпидемических мероприятий
- Дезинфекция 15,2. Дезинфекция (обеззараживание)
- 15.2.2, Дезинсекция
- 15.2.3. Дератизация
- 15.3. Специфическая профилактика инфекционных заболеваний