14 Мочевыделительная система
Заболевания почек и нарушения их функций обычно рассматривают с точки зрения роли этого органа в выведении из организма азотистых шлаков. Ho не менее важна роль почек в поддержании баланса жидкости и электролитов, регуляции АД, кислотно-щелочном гомеостазе. Почка также функционирует как эндокринный орган, гекретируя в кровь гормоны и другие биологически активные вещества (эритропоэтин, простагландины, ренин, активную форму витамина D3).
А. Развитие. Мочевыделительная и половая системы развиваются из промежуточной мезодермы раннего зародыша. При этом последовательно формируются три почки: пронефрос, мезонефрос и метанефрос. Первая из них рудиментарна и не функционирует; вторая действует на ранних стадиях развития плода; метанефрос формирует постоянную почку.
- Пронефрос эмбриона — до 10 скоплений клеток в шейной области. Из них формируются нефротомы (рис. 14-1).
а. Нефротомы. В конце 3-й-начале 4-й недели развития промежуточная мезодерма шейной области отделяется от сомитов и формирует клеточные скопления, имеющие форму стебелька, — нефротомы (рис. 14-1). Нефротомы растут в латеральном направлении, в них образуется полость. Из нефротомов формируются нефрические канальцы, их медиальные концы открываются в полость тела, а латеральные сегменты растут в каудальном направлении. Канальцы соседних сегментов объединяются, формируя парные продольные протоки, растущие по направлению к клоаке (первичный почечный проток). От дорсальной аорты отделяются небольшие ветви, одна из которых внедряется в стенку нефрического канальца, а другая — в стенку целомической полости, формируя соответственно внутренний и наружный клубочки. Клубочки состоят из шаровидного сплетения капилляров и вместе с канальцами формируют выделительные единицы (нефроны). По мере формирования нефротомов пронефроса одновременно происходит их дегенерация, так что в момент образования последнего из них первые нефротомы уже исчезают. К концу 4-й недели развития все признаки нефротомов отсутствуют.
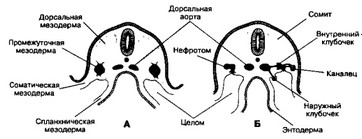
Рис. 14-1. Формирование канальцев наружного и внутреннего клубочков. А — 21-е сутки; Б — 25-е сутки развития (медиальный конец канальца открывается в целом) [из Sadler TW, 1990)
- Мезонефрос (рис. 14-2). По мере дегенерации пронефроса каудальнее появляются первые канальцы мезонефроса. Они удлиняются, формируя S-образную петлю, медиальный конец которой достигает капиллярного клубочка. Последний внедряется в стенку канальца, и в этом месте каналец формирует бдуменову капсулу. Капсула и клубочек образуют почечное тельце. Латеральный конец канальца впадает в первичный почечный проток, который теперь называется мезонефрическим (вольфов). Далее канальцы удлиняются, становясь всё более извитыми. Их окружает сплетение капилляров, образованных постгло- мерулярными сосудами.
а. Гонадный (урогенитальный) валик (рис. 14-3). К середине 2-го месяца мезонефрос достигает максимальной величины; это крупный орган овоидной формы, расположенный по обе стороны от срединной линии. С медиальной его стороны размещается зачаток гонад. Возвышение, формируемое обоими органами, известно как урогенитальный (гонадный) валик. Когда каудальные канальцы ещё дифференцируются, краниальные канальцы и клубочки уже дегенерируют, к концу 2-го месяца большинство из них исчезает. Небольшая часть каудальных канальцев и мезонефрический (вольфов) проток, однако, сохраняются у плода мужского пола. Из канальцев мезонефроса впоследствии формируется рад структур мужской половой системы (см. главу 15.1 Б 2 в (I)).
б. Функция мезонефрических канальцев сходна с функцией канальцев нефрона почки взрослого организма. Фильтрат крови из клубочка поступает в капсулу, затем в каналец и далее — в мезонефрический проток. При этом в канальце происходит реабсорбция ряда веществ. Однако, в мезонефросе моча слабо концентрируется, что связано с отсутствием структур мозгового вещества, необходимых для удержания воды.
Сегментированная
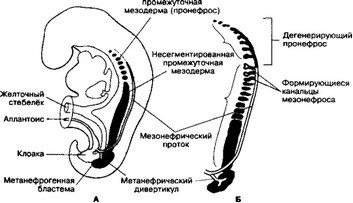
Рис. 14-2. Соотношение промежуточной мезодермы пронефроса, мезонефроса и метанеф- роса. А — 4-недельный эмбрион. В шейной и верхнегрудной областях промежуточная мезодерма сегментирована. В нижнегрудной, поясничной и крестцовой областях промежуточная мезодерма остаётся в виде сплошной массы ткани. Пронефрос формирует продольный проток, переходящий затем в мезонефрический; Б — 5-недельный эмбрион. Образуются мезонефрические канальцы, пронефрос дегенерирует [из Sadler TW, 1990]
- Метанефрос (постоянная почка, рис 14-4) развивается из метанефрогенной бластемы (источник канальцев нефрона) и метанефрического дивертикула (источник собирательных трубочек и более крупных мочевыводящих путей).
а Развитие Метанефрос появляется в течение 5-й недели развития Его канальцы развиваются из метанефрической мезодермы аналогично тому, как это происходило в мезонефросе
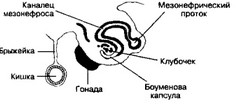 Рис 14-3 Поперечный срез через урогенитальный валик в нижнегрудной области 5-недельного эмбриона Показано формирование канальца мезонефроса, его медиальный конец образует боуменову капсулу, а латеральный впадает в мезонефрический проток На этом сроке развития формируется зачаток гонады Этот зачаток и мезонефрос образуют урогенитальный валик на задней стенке брюшной полости [из Sadler TW1 1990]
Рис 14-3 Поперечный срез через урогенитальный валик в нижнегрудной области 5-недельного эмбриона Показано формирование канальца мезонефроса, его медиальный конец образует боуменову капсулу, а латеральный впадает в мезонефрический проток На этом сроке развития формируется зачаток гонады Этот зачаток и мезонефрос образуют урогенитальный валик на задней стенке брюшной полости [из Sadler TW1 1990]
Задняя кишка
Мезонефрос
Аллантоис
Уроректальная складка
Клоака
Метанефрогенная
бластема
Мезонефрический
проток
Метанефрический
дивертикул
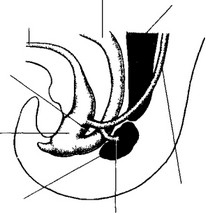
Рис 14-4 Формирование метанефроса (конец 5-й недели развития) Метанефрический дивертикул начинает внедряться в метанефрогенную бластему [из Sadler TW1 1990)
- Метанефрический дивертикул. При впадении в клоаку мезонефрический проток образует вырост — метанефрический дивертикул. Этот вырост внедряется в каудальную часть промежуточной мезодермы, которая уплотняется вокруг дивертикула, образуя метанефрогенную бластему. Далее дивертикул дихотомически делится, формируя систему собирательных протоков, постепенно углубляющихся в ткань метанефроса (рис. 14-5).
- Взаимоиндукции собирательных трубочек и канальцев. Производное метанефрического дивертикула — собирательная трубочка на дистальном конце покрыта «шапочкой» метанефрогенной бластемы. Под индуктивным влиянием трубочек из этой ткани формируются небольшие почечные пузырьки, дающие начало канальцам. В свою очередь развивающиеся канальцы индуцируют дальнейшее ветвление собирательных трубочек.
- Формирование нефронов (рис. 14-6). Канальцы, объединяясь с капиллярным клубочком, формируют нефроны. Проксимальный конец нефрона образует капсулу, в которую глубоко внедряется клубочек. Дистальный конец соединяется с одной из собирательных трубочек. Далее каналец удлиняется, в результате чего образуются проксимальный извитой каналец, петля Хеше и дистальный извитой каналец.
б. Перемещение почки. Сначала почка располагается в области таза. В дальнейшем
она перемещается краниальнее. Кажущийся подъём почки связан с уменьшением
кривизны тела при развитии плода и его ростом в поясничной и крестцовой областях.
в. Функции
- Образование мочи. Моча плода гипотонична относительно плазмы, содержит мало белка и глюкозы, слегка кислая (pH 6,0).
- Поддержание объёма амниотической жидкости — одна из главных функций мочевыделительной системы плода. Начиная примерно с 9-й недели развития, плод выделяет мочу в амниотическую полость (10 мл/кг/час), а также поглощает амниотическую жидкость в объёме до 0,5 л в день.
- Выведение мочевины. Азотистые остатки из организма плода удаляются путём диффузии через плаценту в кровь матери. Следовательно, почки матери служат
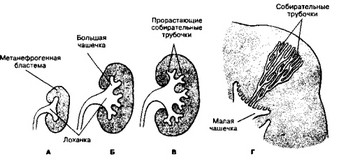
Рис. 14-5. Развитие мочевыводящих путей метанефроса. А — 6-я неделя; Б — конец 6-й недели; В — 7-я неделя развития; Г — у новорождённого. Собирательные трубочки, открываясь в малую чашечку, в совокупности образуют структуру пирамидной формы [из Sadler TW, 1990]
для удаления и материнских, и плодных азотистых остатков. Мочевина легко проходит через плаценту. Часть мочевины попадает в кровь матери, поглощаясь из амниотической жидкости; часть попадает в кровь плода после абсорбции из ЖКТ и затем через плаценту — в кровь матери,
г. Почка новорождённого. У новорождённого почка имеет выраженный дольчатый вид. Дольчатость в дальнейшем исчезает в результате роста, но не формирования de novo нефронов (нефроногенез завершается к 36-й неделе развития, к этому сроку в каждой почке около I млн нефронов).
- Клубочковая фильтрация. При рождении скорость клубочковой фильтрации невелика — около 20-30 мл/1,73 м2/мин; удваивается к двухнедельному возрасту и достигает уровня взрослых к двум годам жизни.
Скорость клубочковой фильтрации (СКФ) определяет количество ультрафильтрата плазмы, образующегося из крови в течение определённого периода времени (в норме СКФ составляет 115-125 мл/мин).
- Реабсорбция натрия. Почки новорождённого не способны к адекватному выделению Na+, что происходит из-за относительной нечувствительности дистального отдела нефрона к альдостерону. Этим также объясняется неполноценность выделения K+ и H+ у новорождённых.
- Способность к концентрации и разведению. Почки новорождённого могут доводить концентрацию мочи до уровня 600-700 мосм/кг (несколько больше половины нормального для взрослых уровня). Способность к разведению достаточна (до 50 мосм/кг), хотя жидкость выделяется медленнее ввиду низкой скорости клубочковой фильтрации.
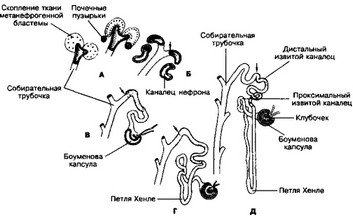
Phс. 14-6. Развитие нефрона. А — 8 недель; Б — 9 недель, В — 11 недель; Г — 20 недель; Д — 25 недель развития. Стрелками показано место перехода канальца нефрона в собирательную трубочку. Сроки указаны приблизительно, т.к. не все нефроны развиваются одновременно [из Johnson KE, 1988]
Способность к концентрации мочи определяют измерением осмоляриости мочи спустя 18-24 часа после прекращения потребления жидкости и повторно после введения 5 ЕД вазопрессина. При таких условиях у 90% здоровых людей осмолярность мочи достигает 900 мосм/кг (или удельного веса 1,023). Способность к разведению мочи определяют измерением осмоляриости мочи и её объёма спустя 5 часов после водной нагрузки из расчёта 20 мл/кг массы тела. У здоровых людей осмолярность мочи достигает 100 моем/кг (или удельного веса 1,003), а объём мочи превышает 80% водной нагрузки.
- Закисление мочи. Снижение реабсорбции гидрокарбоната и секреции ионов H+ приводит к ограничению способности почек к закислению мочи. В результате у новорождённых часто отмечают лёгкий метаболический ацидоз при уровне бикарбоната сыворотки 20-22 мэкв/л.
Способность к образованию кислой мочи pH мочи при голодании в норме ниже 5,5. Способность к образованию кислой мочи оценивают при введении 100 мг хлорида аммония на I кг массы тела с целью уменьшения концентрации бикарбоната плазмы ниже 20 мэкв/л. При этих условиях моча в норме закисляется (те. pH мочи падает ниже 5,5).
- Пороки развития. До половины всех врождённых пороков у детей приходится на долю органов мочеполовой системы.
а. Агенезия (отсутствие) или недоразвитие (гипоплазия) почки наблюдается вследствие ранней дегенерации метанефрического дивертикула или отсутствия индуцирующего влияния со стороны дивертикула на метанефрогенную бластему. Одностороннее отсутствие почки встречается у одного из 1500 новорождённых Двусторонняя гипоплазия наблюдается реже (I из 4000). В этом случае происходит уменьшение количества амниотической жидкости — олигогидрамнион.
б. Эктопия почки — аномалия расположения органа, при этом почка чаще обнаруживается в области таза.
в. Седловидная почка образуется в результате слияния каудальных частей обеих почек.
г. Кистозная болезнь мозгового вещества (нефронофтйз) характеризуется наличием кист в мозговом веществе и атрофией канальцев.
д. Гипоспадйя уретры. Отверстие мочеиспускательного канала открывается на вентральной стороне полового члена.
е Эписпадйи уретры. У мальчиков отверстие мочеиспускательного канала расположено на дорсальной поверхности полового члена. У девочек возможны расщепление клитора либо укорочение мочеиспускательного канала, дефекты шейки мочевого пузыря.
- Наследуемые заболевания почек, как правило, не строго менделирующие. Для некоторых состояний обнаружены типичные сочетания HLA-Ar (табл. 4-4).
а. Наследственный нефрит (синдром длпорта).
б. Поликистозная болезнь почек приводит к прогрессирующей почечной недостаточности. Пересадка почки — единственный эффективный метод лечения.
Б. Почка
- План строения (рис. 14-7). Почка покрыта тонкой капсулой из плотной волокнистой соединительной ткани. Строма почки (интерстиций) — рыхлая волокнистая соединительная ткань, представлена отростчатыми фибробластоподобными клетками и ретикулиновыми волокнами, содержит кровеносные сосуды, в т.ч. вторичную капиллярную сеть, куда из канальцев почки происходит реабсорбция. Почка отчётливо подразделяется на расположенное кнаружи (у выпуклой поверхности органа) корковое вещество и находящееся под ним мозговое вещество.
а. Кора почки содержит почечные тельца и извитые канальцы нефрона.
б. Мозговое вещество. В нём различают наружную часть, расположенную непосредственно под корковым веществом, и внутреннюю часть, состоящую из вершин пира-
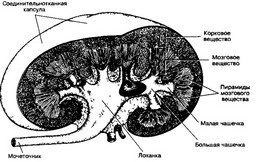
Рис 14 7 Общий плав строения почки Почка покрыта соединительнотканной капсулой, под кото рой расположено корковое вещество (с выпуклой стороны органа) Под корковым веществом находится мозговое, организованное в виде пирамид Основания пирамид обращены к корковому веществу, сосочки открываются в малые чашечки Малые чашечки, сливаясь, образуют большие, открывающиеся в лоханку От лоханки отходит мочеточник [из Kessel RG Kardon RH, 1979]
мид Мозговое вещество почки — уникальная область, характеризующаяся высоким перепадом осмолярности В наиболее глубоких отделах мозговой части почки осмо- лярность в 5 раз превышает осмолярность коры Перепад осмолярности — главная причина реабсорбции воды в Пирамиды мозгового вещества В совокупности собирательные трубочки и прямые участки нефронов образуют пирамиды мозгового вещества, вершины которых обращены к лоханке почки Собирательные трубочки направляются к вершине пирамиды — сосочку, сливаясь друг с другом по мере приближения к последнему, так что калибр трубочек увеличивается и образуется собирательный проток (сосочковый канал), открывающийся на вершине сосочка в полость малых почечных чашечек Последние, сливаясь, образуют большие почечные чашечки, которые представляют собой ответвления расширенного конца мочеточника — почечной лоханки г Доля почки Пирамида мозгового вещества с покрывающей ее основание частью коркового вещества составляет долю почки Почка состоит из 8-18 долей В каждой доле мозговое вещество внедряется в корковое в виде мозговых лучей Центральную часть мозгового луча составляет собирательная трубочка, в которую открываются канальцы многих нефронов, отчего она выглядит ветвящейся Долька мозговой луч вместе с прилежащим кортикальным лабиринтом образует дольку почки
д Кровоток (рис 14-8) В ворота почки входит почечная артерия Она сразу распадается на междолевые артерии, направляющиеся к выпуклой поверхности почки Эти артерии проходят между пирамидами мозгового вещества и могут служить границами долей почки Примерно на границе между корковым и мозговым веществом междолевые артерии поворачивают под прямым углом, образуя дуговые артерии,
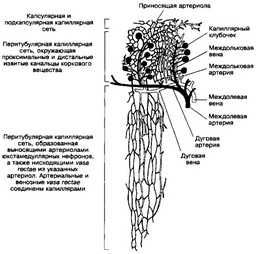
Рис 14-8 Кровоснабжение почки Входящая в почку почечная артерия делится на междолевые, идущие между пирамидами по направлению к выпуклой поверхности почки На границе мозгового вещества с корковым междолевые артерии поворачивают под прямым углом и переходят в дуговые, располагающиеся параллельно выпуклой поверхности почки От дуговых ответвляются междольковые артерии, проходящие между дольками в корковом веществе От междольковых отходят внутри- дольковые (приносящие артериолы), распадающиеся на капилляры, образующие клубочек (первичная капиллярная сеть) Капилляры клубочка собираются в выносящие артериолы, дающие начало вторичной (перитубулярной) капиллярной сети коркового и мозгового вещества, причем перитубу- лярные капилляры глубоких слоев мозгового вещества имеют прямой ход (vasa rectae) Далее начинается венозное русло, ход вен повторяет ход одноименных артерий [из Kessel RG, Kardon RH, 1979]
располагающиеся в плоскости, параллельной поверхности почки. От дуговых артерий отходят, снова направляясь к выпуклой поверхности, междольковые артерии. Эти артерии определяют границы долек почки в корковом веществе, центральную часть которых составляют мозговые лучи. Любой отдел артериального русла почки не имеет анастомозов между собой. Следовательно, прекращение кровотока в артериальном русле приводит к гибели той части паренхимы почки, которая получает кровоснабжение от данного отдела артериального русла. Ход венозных сосудов практически повторяет ход артериальных.
- Первичная капиллярная сеть. От междольковых артерий параллельно поверхности органа ответвляются короткие приносящие артериолы (внутридолько- вые артериолы), переходящие в капиллярный клубочек (первичная капиллярная сеть). Клубочки первичной капиллярной сети входят в состав почечных телец, в которых происходят фильтрация плазмы и образование клубочкового фильтрата. Выносящая артериола собирает кровь из капилляров клубочка.
- Вторичная капиллярная сеть — вторая капиллярная сеть почки, в которую кровь поступает из первичной капиллярной сети через выносящие артериолы. Эти артериолы переходят в прямые сосуды, залегающие в мозговом веществе. Они проходят параллельно канальцам нефронов и собирательным трубочкам, отчего и получили название vasa rectae. Прямые сосуды образуют капилляры вторичной сети, оплетающие канальцы петли Хёнле. Капилляры вторичной (периту- булярной) сети располагаются в непосредственной близости от канальцев нефронов; в эти капилляры осуществляется реабсорбция веществ из просвета канальцев. Из вторичной капиллярной сети также происходит питание ткани почки. Кровь из капилляров мозгового вещества поступает в прямые венулы, впадающие в дуговые вены.
- Нефрон (рис. 14-9). Паренхима почки состоит из 1-2 миллионов функциональных структурных единиц — нефронов, а также системы собирательных протоков. Извитые части нефронов располагаются в корковом веществе, тогда как их прямые участки — в мозговом. Извитые канальцы нефронов составляют кортикальный лабиринт. Дистальный каналец каждого нефрона открывается в собирательную трубочку. Нефрон — эпителиальная трубка, начинающаяся от почечного тельца и впадающая в собирательную трубочку. Стенка нефрона построена из однослойного эпителия, клетки которого (в зависимости от выполняемой функции) различны в разных отделах нефрона.
а. Отделы. В нефроне различают несколько отделов: капсула почечного тельца, в состав которого входит капиллярный клубочек; проксимальный извитой и проксимальный прямой канальцы, тонкий каналец; дистальный прямой и дистальный извитой канальцы. Тонкий и дистальный прямой канальцы образуют нисходящую и восходящую части петли Хёнле. Дистальный прямой каналец возвращается к собственному почечному тельцу и контактирует с ним; далее он переходит в дистальный извитой, впадающий в собирательную трубочку.
Локализация. Разные отделы нефрона закономерно расположены либо в корковом, либо в мозговом веществе. В корковом веществе располагаются: почечное тельце, состоящее из капиллярного клубочка и его капсулы, проксимальный и дистальный извитые канальцы, окружённые капиллярами вторичной капиллярной сети. В мозговом веществу располагается петля Хёнле, также сопровождаемая капиллярами. Тут же присутствуют и собирательные трубочки.
б. Типы нефронов. Различают два основных типа нефронов — кортикальный и юкстамедуллярный. 85% всех нефронов — кортикальные, юкстамедуллярных — 15%.
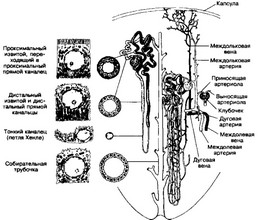
Рис. 14-9. Нефрои. Строение отделов и связь с кровеносными сосудами. От капсулы клубочка (капилляры первичной сети) начинается проксимальный извитой каналец, переходящий в проксимальный прямой. Оба отдела нефрона состоят из высокого кубического каёмчатого эпителия, клетки которого имеют выраженную базальную исчерченность и много митохондрий Дистальные канальцы (прямой и извитой) образованы кубическими клетками с базальной исчерченностью и малым количеством микроворсинок. Тонкий каналец имеет узкий просвет и состоит из уплощённых эпителиальных клеток. Канальцы нефрона оплетены капиллярами перитубулярной (вторичной) сети, которые дают начало венозному руслу. Собирательные трубочки имеют широкий просвет и образованы кубическим эпителием [из Hees Н, Sinowatz F1 1992; Junqueira LC, Cameiro J, 1991]
- Кортикальный нефрон. У кортикального нефрона почечное тельце расположено в наружной части коркового вещества. Петля Хеше у большей части кортикальных нефронов имеет небольшую длину и располагается в пределах наружной части мозгового вещества.
- Юкстамедуллярный нефрон. Почечное тельце расположено на границе с мозговым веществом. У большинства юкстамедуллярных нефронов длинная петля Хенле проникает глубоко в мозговое вещество, вплоть до верхушек пирамид.
в. Почечное тельце (рис. 14-10) состоит из капиллярного клубочка и его эпителиальной капсулы, являющейся началом нефрона. Здесь происходят фильтрация и образование первичной мочи (10% объёма крови, протекающей по капиллярам клубоч-
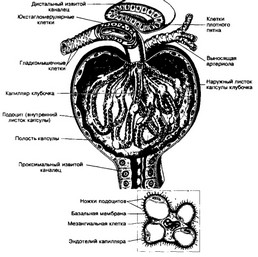
Рис 14 10 Почечное тельце На рисунке внизу показана локализация мезангиальных клеток между капиллярными петлями клубочка [из Hees Н, Smowatz F, 1992]
ка, ультрафильтрат составляет примерно 180 л/сутки) Приносящая артериола распадается примерно на 50 капиллярных петель, собирающихся в выносящую артериолу Область, где в тельце входит приносящая и выходит выносящая артериолы, называют сосудистым полюсом
- Капсула клубочка состоит из двух листков наружного (париетального) и внутреннего (висцерального) Между листками имеется полость, куда из просвета кровеносных капилляров поступает клубочковый фильтрат Полость капсулы открывается в проксимальный извитой каналец
(а) Наружный листок капсулы, состоящий из однослойного плоского эпителия, ограничивает капсулярное пространство снаружи
(б) Внутренний листок капсулы (рис 14-11) Клетки внутреннего листка капсулы (подоциты) прикреплены к наружной поверхности капилляров клубочка и вместе с эндотелием и базальной мембраной, общей для капилляра и подо- цитов, участвуют в процессе фильтрации Подоциты содержат ядро неправильной формы с глубокими инвагинациями, хорошо развитый комплекс Гольджи, микротрубочки и филаменты, которых особенно много в отростках клеток От тела подоцита отходят ветвящиеся отростки, или ножки Отростки ветвятся, в результате на поверхности капилляра образуется лабиринт из щелей между ножками подоцитов, прикрепляющихся к базальной мембране — фильтрационные щели (рис 14-12) Они имеют ширину около 30—40 нм и затянуты диафрагмами (сеть из волокнистых структур) Размеры ячеек сети — около 10 нм Ножки подоцитов в широких пределах изменяют свой диаметр, что неизбежно сказывается на ширине фильтрационных щелей Эффективный диаметр фильтрационных щелей таков, что через них могут проходить молекулы веществ с массой не более 50 кД
- Фильтрационный барьер состоит из эндотелия капилляров, базальной мембраны и щелевых диафрагм между ножками подоцитов Фильтрационные щели — главная часть барьера Поток жидкости сквозь барьер обеспечивается ее гидростатическим давлением Это давление понижается онкотическим давлением белков плазмы
(а) Эндотелиальные клетки капилляров клубочка максимально уплощены, за исключением области, содержащей ядро Тонкая часть клетки пронизана фенест- рами (порами) полигональной формы диаметром 50-100 нм, занимающими примерно 30% всей поверхности клетки В результате плазма крови непосредственно контактирует с базальной мембраной
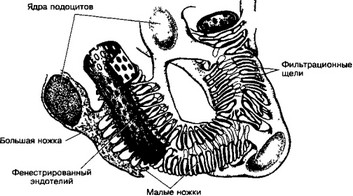
Рис 14-11 Связь подоцитов с эндотелием капилляров Подоциты — видоизмененные эпителиальные клетки внутреннего листка капсулы Они образуют большие ножки, от которых отходят многочисленные нитевидные малые ножки Уплощенные эндотелиальные клетки капилляров клубочка имеют многочисленные фенестры Между внутренним листком капсулы и эндотелием капилляров формируется общая базальная мембрана [из Ham А, 1974]
- Базальная мембрана толщиной до 300 нм формируется (в основном за счет синтетической деятельности подоцитов) как непрерывный слой, общий для подоцитов и эндотелия капилляров В базальной мембране различают три слоя, средний из которых — наиболее толстый и электроноплотный, а наруж ный и внутренний слои представляются более разреженными Базальная мембрана образована ворсистой структурой тонких (около 2-5 нм) филаментов (коллаген IV типа) в виде молекулярного сита, филаменты погружены в гомогенный
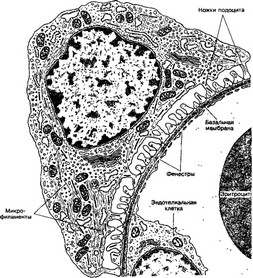
Рис 14-12 Фильтрационный барьер Малые ножки подоцитов прикрепляются к трехслойной базальной мембране, общей для подоцитов и эндотелия капилляров клубочка Между ножками подоцитов имеются узкие (30-40 нм) фильтрационные щели Фильтрация плазмы осуществляется через волокнистую основу базальной мембраны и фильтрационные щели [из Lentz TL, 1971]
гликопротеиновый матрикс. Вещества с Mr до 10 кД проходят через это сито свободно, а более 50 кД — в ничтожных количествах. На основании этих данных максимальный размер пор базальной мембраны оценивается в 4 нм.
- Мезангиальные клетки. Внутренний листок капсулы не покрывает полностью каждый отдельный капилляр клубочка. Между капиллярами, не имеющими в таких местах общей с эпителием базальной мембраны, располагаются клетки отрост- чатой формы — мезангиальные клетки.
(а) Рецепторы. Мезангиальные клетки имеют рецепторы ангиотензина II, атри- опептина и вазопрессина. Вазопрессин стимулирует сокращение мезангиаль- ных клеток.
(б) Функции
- Сократительная активность. В цитоплазме мезангиальных клеток в большом количестве присутствуют микрофиламенты; вероятно, эти клетки обладают сократительной активностью. Благодаря этому, клетки способны уменьшать площадь поверхности стенки капилляров, через которую происходит фильтрация, уменьшая таким образом уровень последней.
- Фагоцитоз. Мезангиальные клетки способны фагоцитировать остатки базальных мембран.
- Синтез макромолекул
- Мезангиальные клетки синтезируют макромолекулы межклеточного вещества, присутствующего между капиллярами.
- Мезангиальные клетки синтезируют фактор активации тромбоцитов (PAF).
г. Канальцы нефрона (рис. 14-9) и реабсорбция. Каналец нефрона начинается от
почечного тельца и содержит в просвете первичную мочу, поступающую сюда из полости капсулы тельца. Каналец нефрона имеет извитой ход и разную толщину стенки за счёт различной высоты эпителиальных клеток. В разных отделах канальца эпителиальные клетки имеют особенности строения в зависимости от того, что именно реабсорбируется в данном отделе. То, что подлежит экскреции, выбросу из организма (например, мочевина, мочевая кислота, креатинин), не реабсорбируется. Наконец, через клетки канальцев происходит не только реабсорбция, но и движение в обратном направлении, т.е. из кровеносных капилляров в просвет канальца (секреция). Суточный объём реабсорбции в канальцах нефрона и собирательных трубочках приближается к объёму первичной мочи (за вычетом, естественно, дефинитивной [вторичной! мочи).
- Общие свойства эпителия. Характерны цитоплазматические отростки на базальной и латеральных поверхностях клеток и интердигитации с соседними клетками. В апикальных частях соседние клетки образуют между собой обширные плотные контакты. Ниже области контактов клетки отделены друг от друга латеральным межклеточным пространством. Это пространство функционально продолжается в окружающее каналец интерстициальное (перитубулярное) пространство. Базальная мембрана эпителия не препятствует транспорту веществ в эти пространства.
- Реабсорбция (рис. 14-13) различных веществ осуществляется путём активного транспорта, тогда как вода выходит из канальцев пассивно. При этом имеет значение разница в осмотическом давлении между просветом канальцев и окружающими каналец структурами, т.е. капиллярами вторичной сети, а также разница в осмотическом давлении между корковым и мозговым веществом.
(а) Ионы и вода. Активно реабсорбируются ионы (например, ионы Na+), но не везде, а в проксимальном канальце, в дистальном канальце, в толстом отделе петли Хёнле. Эти структуры расположены либо в глубоких слоях коркового,
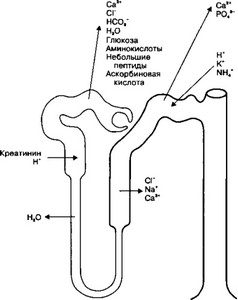
Рис 14-13 Функции канальцев нефрона В отделах нефрона происходит реабсорбция различных веществ из первичной мочи Основное количество ионов (Na+, Cl', HCO3', Ca2+) реабсорбируется в проксимальных и дистальных канальцах Глюкоза, аминокислоты, небольшие белковые молекулы полностью реабсорбируются в проксимальном извитом канальце Креатинин секретируется в просвет проксимального канальца Ионы К*, Ht, NH4* поступают в основном в дистальный каналец. Вода пассивно реабсорбируется в проксимальном канальце, тонком отделе петли, в собирательных трубочках
либо в поверхностных слоях мозгового вещества. По мере продвижения по канальцу его содержимое из гипертонического становится гипотоническим, а среда вне канальцев по мере продвижения от коркового к мозговому веществу становится всё более гипертоничной. В силу разницы давлений вода будет выходить из просвета канальцев в окружающие их кровеносные капилляры, что и происходит в тонком отделе петли Хёнле и в собирательных трубочках, т.е. структурах, расположенных в мозговом веществе. Такой пассивный транспорт воды тем больше, чем больше разница в осмотическом давлении, т.е. чем глубже петля нефрона внедряется в мозговое вещество.
(б) Активный транспорт. В клетках канальцев нефрона, активно транспортирующих ионы Na+, №+,К+-АТФаза расположена преимущественно в базальной части клетки и отчасти — на её боковых поверхностях; здесь же сосредоточены и митохондрии. Поступивший по электрохимическому градиенту из просвета
канальца в клетку Na+ активно транспортируется в латеральное межклеточное пространство, далее — в перитубулярное, а затем — в расположенный рядом с канальцем капилляр.
(в) Транспорт Na+ осуществляется путём обмена внеклеточного Na+ на внутриклеточный H+ при помощи белков плазмолеммы NHE.
- NHEl и NHE2 экспрессируются в базолатеральных отделах плазмолеммы эпителия канальцев почки. Диуретик амилорид ингибирует NHEl и NHE2. Дефекты гена SLC9A1 приводят к артериальной гипертензии (эссенциальная гипертоническая болезнь).
- NHE3 апикальной мембраны эпителиальных клеток канальцев почки важен для трансэпителиального переноса Na+ и не чувствителен к амилориду.
д. Проксимальный каналец. Стенка канальца образована кубическим эпителием. На боковых поверхностях клетки имеются складки, так что между соседними клетками образуются интердигитации. Между клетками канальца в апикальной части формируются плотные контакты, отделяющие межклеточное пространство от просвета канальца. Из просвета проксимальных извитых канальцев в окружающие их кровеносные капилляры перекачивается 80% ионов натрия и хлора, а также вода, практически вся глюкоза и весь отфильтрованный в почечных тельцах белок. В проксимальном канальце в его просвет секретируются лекарственные препараты и их метаболиты, креатинин.
- Эпителий. Для клеток характерно крупное округлое ядро, множество пиноцитоз- ных пузырьков, вакуолей и масса лизосом.
(а) Апикальная часть. На апикальной поверхности клетки имеются многочисленные микроворсинки, образующие щёточную каёмку. На микроворсинках выявлена высокая активность щелочной фосфатазы. Между основаниями микроворсинок отходят трубки — апикальные (верхушечные) канальцы.
(б) Базальная часть. Мембрана базальной части формирует глубокие впячива- ния, разделяющие эту часть клетки на переплетающиеся между собой и с аналогичными структурами соседних клеток отростки.
(в) Лизосомы. При помощи лизосом в эпителии канальцев расщепляется захваченный материал, в первую очередь белок. Молекулы белка попадают в апикальные канальцы, которые отрываются от плазмолеммы с образованием апикальных пузырьков. Далее апикальные пузырьки сливаются с лизосомами, и полипептиды расщепляются до аминокислот. Затем аминокислоты переносятся в кровеносный капилляр через боковые поверхности клетки и через её базальную часть.
(г) Митохондрии. Во всем объёме клетки, за исключением её апикальной части, присутствует множество митохондрий, ориентированных вдоль длинной оси клетки.
- Прямой отдел. В прямом отделе проксимальный каналец имеет в целом такую же структуру, что и в извитом. Клетки в этом отделе нефрона ниже, имеют меньше боковых складок. Количество микроворсинок постепенно снижается по мере приближения к тонкому отделу.
- Реабсорбция
(а) Фосфаты. Проксимальные канальцы — главный участок реабсорбции фосфатов. Фосфат реабсорбируется в сочетании с транспортом Na*. Реабсорбция фосфатов происходит при помощи кодируемых генами NPT2 переносчиков (глава 2 I В I б (I) (г) (i)].
Дефекты генов NPT2 приводят к почечным потерям фосфатов при ряде наследуемых заболеваний (гипофосфатемия, X; гипофосфатемический рахит, SR, р; гипофосфатеми- ческий рахит с гипокальциурией, гипофосфатемическая болезнь костей).
(б) Глюкоза реабсорбируется путём сочетанного транспорта с Na+ при помощи мембранных гликопротеинов, кодируемых генами SGLT. Это главный механизм почечной реабсорбции глюкозы. Экскреция глюкозы с мочой в норме обычно не превышает 0,3 г/сутки.
Мутации гена SGLT2 приводят к глюкозурии — почечной потере глюкозы (до 60 г в сутки).
(в) Аминокислоты. Реабсорбция аминокислот осуществляется при помощи переносчиков, кодируемых генами SLC.
Цистинурия Мутации гена SLC3A1 приводят к избыточной секреции цистина (при некоторых формах также лизина, аргинина и орнитина) и к образованию мочевых камней
(г) Белок. Реабсорбция белка из просвета канальцев происходит путём пиноци- тоза. Молекулы белка расщепляются до аминокислот при помощи многочисленных лизосом. Образовавшиеся аминокислоты транспортируются с участием специальных переносчиков в капилляры вторичной сети. Для у
Источник: Под ред. Э.Г. Улумбекова, Ю.А. Челышева, «Гистология (введение в патологию)» 1997
А так же в разделе «14 Мочевыделительная система »
- ИСТОЧНИКИ И РАЗВИТИЕ
- ПЛАН СТРОЕНИЯ ПИЩЕВАРИТЕЛЬНОГО ТРАКТА
- НЕРВНЫЙ АППАРАТ
- ЭНДОКРИННАЯ СИСТЕМА
- V РОТОВАЯ ПОЛОСТЬ
- ПИЩЕВОД
- ЖЕЛУДОК
- IX ТОНКИЙ КИШЕЧНИК
- ТОЛСТЫЙ КИШЕЧНИК
- ЖЕЛЕЗЫ ПИЩЕВАРИТЕЛЬНОГО ТРАКТА
- ПРЕПАРАТЫ
- ОТВЕТЫ И ПОЯСНЕНИЯ
- 13 Дыхательная система
- ПРЕПАРАТЫ
- ОТВЕТЫ И ПОЯСНЕНИЯ
- ПРЕПАРАТЫ
- ОТВЕТЫ И ПОЯСНЕНИЯ