Внешнее (латеральное) торможение
Внешнее (латеральное) торможение выученной реакции наблюдается при предъявлении незнакомого стимула достаточной значимости и интенсивности. Мы попытаемся объяснить это явление в рамках предлагаемой модели. Для этого потребуются дополнительные комментарии по поводу роли ГМЛ.
Какизвестно, стриосомы посылают эфферентные проекции в черную субстанцию (pars compacta), которая в свою очередь дает дофаминергические проекции в моторные области стриатума (рис. 6.8). Мы предполагаем, что стимул, вызывающий внешнее торможение, как и любой другой незнакомый стимул, активирует ГМЛ, снижает дофаминергичес- кую иннервацию моторных отделов стриатума и тем самым нарушает воспроизведение выученной поведенческой последовательности. Такое объяснение согласуется с традиционными взглядами на механизмы внешнего торможения (Mackintosh, 1974, с. 16).
Недавно было высказано предположение, что одной из функций стриатума является "разбиение" поведения на модули, состоящие из отдельных элементов, актов, поз (Graybiel, 1998; см. также Pellis et al., 1993). Вероятно, инициация и автоматическое воспроизведение каждого модуля поведения опосредованы различными отделами стриатума. Это предположение подтверждается разделением стриатума на лимбические (вентральные) и моторные (дорсальные) отделы. Предлагаемая модель объясняет
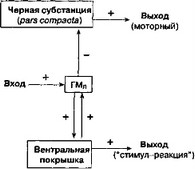
Рис. 6.8. Схематическое представление механизмов, лежащих в основе внешнего торможения (объяснение в тексте)
- Регуляция активности гностических модулей
работу вентральных отделов стриатума. Дорсальные отделы обеспечивают автоматическое исполнение поведенческих модулей. Соответственно, вентральные и дорсальные отделы стриатума иннервируются различными популяциями мезэнцефальных дофаминергических нейронов (ВТО и ЧС). Функция стриатума, связанная с "разбиением" поведения на отдельные модули, призвана разрешить трудности, возникающие, когда механистическая нейробиологическая или поведенческая модель используется для анализа единичных стимулов и реакций, искусственно выхваченных из контекста более или менее постоянного "потока" поведения (Schoenfeld, 1972).
Растормаживание—еще одно явление, которое, возможно, тесно связано с внешним торможением. Когда внешний стимул предъявляют во время сеанса угашения ранее выученной поведенческой реакции, амплитуда этой реакции может достоверно возрасти. Растормаживание также наблюдается в процессе привыкания (габитуации) и критически зависит от интенсивности и новизны "растормаживающих" стимулов (Mackintosh, 1974, с. 16, 38). В рамках предлагаемой модели феномен растормажива- ния следует рассматривать как особый случай взаимодействия двух альтернативных связей "стимул-реакция" — той, которая ранее была выучена, и другой, которая соответствует врожденной ориентировочной реакции на новый стимул (см. выше).
Источник: Беспалов А. Ю., Звартау Э. Э., «Нейропсихофармакология антагонистов NMDA-рецепторов» 2000
А так же в разделе «Внешнее (латеральное) торможение »
- 5.2.1. Реакция внутривенного самовведения
- Реакция электрической самостимуляции мозга
- 5.3.1. Реакция условнорефлекторного предпочтения места
- Реакция электрической самостимуляции мозга
- Условнорефлекторное восстановление угашенной реакции внутривенного самовведения
- Условнорефлекторная локомоторная активность
- Сенситизация (обратная толерантность)
- Теоретическое обоснование влияния антагонистов NMDA- рецепторов на подкрепляющие свойства наркотиков
- Стриарные механизмы памяти и обучения: поискуниверсального механизма подкрепления на поведенческом уровне
- Принцип образования связи "стимул-реакция"
- Долговременные изменения в глутаматергических синапсах стриатума
- Г ностические модули и феномен "перепрограммирования" стриарных проекций
- Взаимодействие дофамина и глутамата
- Лимбико-моторная интеграция и проблема частного и целого
- Ожидание "награды" и "запускающий механизм"
- "Запускающий механизм": дофамин или глутамат?
- Регуляция "запускающего механизма"
- Альтернативное подкрепление и адъюнктивное поведение
- Новизна стимулов
- Основные поведенческие феномены в свете предлагаемой модели
- Угашение
- . Латентное торможение
- Избирательность образования связей "стимул-реакция"
- Регуляция активности гностических модулей
- Идеальная экспериментальная модель для анализа вклада глутаматергической системы в