Серотонинергическая система
Серотонин, или 5-гидрокситриптамин (5НТ), как норадреналин и Дофамин, относится к моноаминам. Источник его синтеза — аминокислота триптофан, поступающая в организм с пищей, где под влиянием ферментов из него образуется 5-гидрокситриптофан (-5НТР), а затем 5-гидрокситриптамин, т.е. серотонин. Самая медленная фаза синтеза серотонина — превращение триптофана в 5 'гидрокситриптофан. Именно эта ступень определяет время течения всего синтеза. Синтезированный в синаптических окончаниях серотонин с помощью высокоаффинных переносчиков депонируется в везикулах. После выхода в синаптическую щель ( экзоцитоз) часть серотонина взаимодействует с рецепторами,
65
а его излишки удаляются из синаптической щели системой активного обратного захвата. Разрушение серотонина в тканях происходит под влиянием моноаминоксидазы. В серотонинергичес- ких нейронах обнаружена оксидаза типа МАОа.
Сходство между двумя биогенными аминами — норадреналином и серотонином — проявляется и в одинаковом действии на них некоторых нейротропных веществ. Так, резерпин вызывает уменьшение содержания в мозге как норадреналина и дофамина, так и серотонина.
Серотонинергические нейроны и пути. Тела серотонинергических нейронов формируют обособленные ядра, так называемые ядра шва (Bt-Bg). «Верхняя» группа этих ядер (В5-В9) находится в ростральной части моста мозга (рис. 13). Их нейроны дают проекции рострально, в основном ипсилатерально. Наиболее важный тракт, в составе которого идут миелинезированные и немиелинезированные волокна серотонинергических нейронов, — медиальный пучок переднего мозга. Эти волокна иннервируют в основном лимбическую систему, базальные ганглии, кору больших полушарий. Обширно разветвляясь, они образуют перекрывающиеся терминальные поля. Однако есть свидетельства о некоторой топографической организации проекций нейронов ядер шва. Так, волокна, выходящие из дорзального ядра шва, образуют серотонинергические проекции преимущественно в базальных ганглиях и в мозжечке, тогда как волокна медиального ядра шва обеспечивают большое количество входов в гиппокамп и септум.
В продолговатом мозге находятся серотонинергические нейроны, формирующую «нижнюю» группу ядер (Bt-B4). Их аксоны образуют нисходящие пути в ствол мозга и в спинной мозг (см. рис. 13). Эти нейроны играют ключевую роль в регуляции двигательной активности, вегетативных функций, регуляции проведения болевой чувствительности.
Рецепторы. В серотонинергической системе на основе структуры и влияния фармакологических веществ выделяют 7 типов серотонино- вых рецепторов (5НТ,, 5НТ2,5НТ3 и т.д.), внутри каждого типа — подтипы (около 15). Все серотониновые рецепторы относятся к метаботропным рецепторам. 5НТ,-рецепторы (подтипы А, В, D, Е) сопряжены с аденилатциклазой (влияют на синтез цАМФ); 5НТ2-рецепторы (подтипы А, В, С) — с фосфолипазой, с образованием вторичных мессенджеров диацилглицерола и трифосфоинозитола; серотониновые рецепторы 5НТ4,5НТ6 и 5НТ7 — с С5-белком (при их взаимодействии
Гиппокамп Гиппокамп Таламус Мозжечок
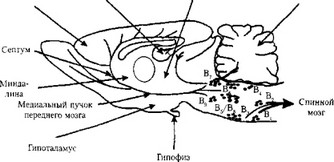
Рис. 13. Серотонинергические ядра шва и структуры мозга, в которые приецируются серотонинергические волокна (цит. по: Stanford S. С., 2001):
В,-В4 — «нижняя» группа ядер шва; В5-В9 — «верхняя» группа ядер шва
с медиатором активируются аденилатциклаза и цАМФ-зависимые протеинкиназы, а также К+- и Са2+-каналы).
Серотониновые рецепторы широко распространены в мозге. Высокая концентрация 5НТ 1А-рецепторов обнаружена в гиппокампе, сеп- туме, миндалине, на телах серотонинергических нейронов ядер шва. Их активация сопровождается снижением синтеза цАМФ, что ведет к увеличению К+-тока, а следовательно, к гиперполяризации мембраны, торможению активности клеток и к снижению высвобождения серотонина из терминалей нейронов ядер шва. Активация 5НТ1А-рецепто- ров приводит к уменьшению тревоги, а известный агонист этих рецепторов, буспирон, используется в клинике в качестве анксиолитика.
5НТ1В-рецепторы присутствуют в базальных ганглиях, стриату- ме, фронтальной коре и пресинаптически на терминалях серотонинергических нейронов. Их возбуждение приводит к уменьшенному высвобождению 5НТ, однако существуют некоторые региональные различия в эффектах активации. Полагают, что эти рецепторы участвуют в регуляции циркадного ритма (уменьшая высвобождение серотонина в супрахиазматическом ядре гипоталамуса) и двигательной активности (увеличивая ее при активации). Получены также данные, что 5НТ1В-рецепторы регулируют высвобождение других нейротрансмиттеров из терминальных окончаний, в частности, дофамина, норадреналина, глутамата.
5Ш,грецепторы локализованы преимущественно в коре и базальных ганглиях. Их активация приводит к уменьшению ^-проводимости и возбуждению нейронов. Известные агонисты этих рецепторов — LSD (диэтиламид лизергиновой кислоты) и DOM (2,5-диметокси-4-метиламфетамин) — относятся к галлюциногенам. Многие атипичные нейролептики (азалепин, рисперидон, клоза- пик) - антагонисты этих рецепторов.
5Шх-рецепторы находятся в гипоталамусе, септуме, миндалине и мозжечке. Интерес к ним вызван тем, что некоторые их агонисты (м-хлорфенилпиперазин) проявляют анксиогенную активность и снижают потребность в пище.
5НТ^рецепторы (в коре головного мозга, стволе, миндалине, гиппокампе) хорошо известны тем, что их активация способствует высвобождению из терминали дофамина и других нейромедиаторов, включая ГАМК, ацетилхолин, норадреналин. Агонисты 5НТ^рецепторе способствуют активации когнитивных процессов в ЦНС.
5НТ-рецепторы обнаружены в коре, миндалине, аккумбентном ядре, стриатуме. Многие антипсихотические вещества и некоторые антидепрессанты проявляют высокую аффинность к ним, выступая их антагонистами. Плотность 5НТ?-рецепторов наиболее высока в таламусе, гиппокампе и гипоталамусе (супрахиазматическом ядре). Как полагают, они принимают участие в синхронизации циркадных ритмов со световым циклом дня и ночи.
Функции серотонинергической системы в ЦНС включают: регуляцию цикла сна и бодрствования, настроения, эмоционального поведения, памяти, аппетита, возбудимости мотонейронов, регуляцию проведения сенсорных стимулов (в том числе и болевых), центральную терморегуляцию, влияние на продукцию ряда гипоталамических факторов и гипофизарных гормонов (кортикотропин-рилизинг фактора, адренокортикотропного гормона).
Регуляция цикла сон—бодрствование. Еще в 1974 г. М. Жюве (Jouvet) показал, что увеличение серотонинергической передачи способствует наступлению сна. В опытах разрушение у кошки ядер шва или блокада синтеза серотонина устраняли синхронизацию ЭЭГ и медленноволновый сон. Однако через несколько недель после поражения ядер шва сон нормализовался, что не подтверждает исключительную роль серотонина в медленноволновом сне.
Прямая стимуляция ядер шва или введение предшественника серотонина увеличивали длительность бодрствования, что предполагает возбуждающее влияние 5НТ на поведение. В электрофизиологических экспериментах активность серотонинергических нейронов дорсального ядра шва в фазе медленноволнового сна становилась менее регулярной и прогрессивно снижалась по сравнению с бодрствованием. Эти нейроны были максимально активны во время бодрствования, но оставались «молчащими» в стадии парадоксального сна. Более того, увеличение скорости разрядов нейронов дорсального ядра шва предшествовало повышению уровня arousal. Полагают, что серотонинергическая передача оказывает модулирующее влияние на характер ответов целевых клеток, регулируя уровень их возбуждения в соответствии с общим уровнем активации ЦНС.
Большое разнообразие серотониновых рецепторов объясняет парадоксальность роли серотонина в регуляции цикла сон—бодрствование. Так, активность 5НТ1А-, 5НТ1В-, 5НТ2А/С- и 5НТ?-рецеп- торов связывают с регуляцией циркадных ритмов. Активация пре- синаптических 5НТ1В-рецепторов в ретиноталамическом тракте уменьшает высвобождение серотонина, ограничивая зрительные входы в латеральные коленчатые тела. Активация постсинаптических 5НТ2С-, 5НТ7- и, возможно, 5НТ)Д-рецепторов, напротив, облегчает зрительную активацию латеральных коленчатых тел, вызывая фазовые сдвиги циркадных ритмов.
Серотониновые рецепторы могут непосредственно участвовать в регуляции сна. Возбуждение постсинаптических 5НТ)Д-рецепто- ров в базальных ганглиях введением их агонистов способствует увеличению длительности медленноволнового сна. Активация преси- наптических 5НТ1Д-рецепторов в холинергических структурах ствола мозга уменьшает активность холинергических нейронов, в результате уменьшается парадоксальная фаза сна. Агонисты 5НТ2Д/с-рецепторов увеличивают время бодрствования и уменьшают медленноволновый и REM-сон у человека и крысы, оказывая влияние, как полагают, на таламические структуры. Введение агонистов 5НТ3-рецепторов в аккумбентное ядро увеличивает бодрствование, уменьшает медленноволновый сон, однако не влияет на парадоксальный сон.
Регуляция пищевого поведения. В этом процессе участвуют многие нейромедиаторы и гормоны, однако ключевая роль принадлежит серотонинергической системе. В экспериментах увеличение серото- нинергической передачи в мозге вызывало уменьшение потребления пищи. Введение 5НТ в паравентрикулярное ядро гипоталамуса крысы приводило к насыщению животного, тогда как при потреблении пищи отмечается усиленный выход серотонина в латеральном гипоталамусе. Полагают, что эти два отдела гипоталамуса выполняют противоположную функцию в регуляции аппетита: недостаточное торможение серотонинергической передачи в латеральном гипоталамусе может быть причиной избыточного потребления пищи при ожирении, а усиленный выход серотонина в паравентрикуляр- ном ядре гипоталамуса — способствовать вызванной стрессом гипо- фагии.
Регуляция эмоционального состояния. В дорсальном и медиальном ядрах шва обнаружены популяции клеток, которые в отличие от большинства других нейронов не реагируют на изменение состояния бодрствования, но отвечают на действие аверсивного (отрицательного) стимула. Однако поведенческие проявления изменения активности серотонинергической медиации и содержания серотонина в мозге весьма противоречивы. Например, увеличенный выход серотонина отмечался в переднем мозге крыс, когда их подвергали неизбегаемому, неконтролируемому стрессу (у животных развивалась беспомощность), а также в состоянии тревоги при действии условного сигнала, предшествовавшего электрокожному подкреплению. С другой стороны, при повышении уровня серотонина в головном мозге животных, которое достигалось введением его предшественника L-триптофана, эмоционально-стрессовая реакция в период прагматического дефицита информации на начальных этапах выработки условного рефлекса уменьшалась. Исчезали характерные для начального периода обучения хаотичность и избыточность двигательной активности, поведение становилось более целесообразным, уменьшался страх. При накоплении серотонина в головном мозге нарушалась выработка условной реакции активного избегания, что, видимо, связано с ослаблением подкрепляющего действия болевого раздражения. Полагают, что эндогенно накопленный серотонин оптимизирует поведение животных в состоянии эмоционального стресса.
Снижение уровня серотонина в мозге кошек и крыс при хроническом введении парахлорфенилаланина приводило к нарушению поведения — развитию значительной сексуальной активности, агрессивности поведения, улучшению выработки условного рефлекса с отрицательным подкреплением. Увеличение агрессивности поведения крыс наблюдалось также при повышении уровня катехоламинов в мозге. Результаты этих экспериментов подтверждают предположение об антагонистических взаимоотношениях, существующих между катехоламинами и серотонином в регуляции агрессивного поведения.
Многие данные свидетельствуют о важной роли биогенных аминов (серотонина, дофамина и норадреналина) в развитии патологии настроения, аффектов. Существует точка зрения, что с ростом концентрации серотонина в мозге настроение у человека поднимается, а его истощение вызывает состояние депрессии. Причем дефицит норадреналина проявляется депрессией тоски, а серотонина — депрессией тревоги. Результаты обследования мозга больных, покончивших с собой в состоянии депрессии, показали, что он обеднен как норадреналином, так и серотонином. Полагают, что дефицит или ингибирование 5НТ6-рецепторов связано с депрессивным состоянием, а их активация — с галлюцинациями.
Некоторые формы фобий также связаны с изменением уровня серотонина, что свидетельствует о важной роли серотонинергической системы в генерации психотических синдромов. При лечении психозов эффективными оказались атипичные нейролептики — антагонисты не только Д2-, но и 5НТ2Д-рецепторов, они не вызывают экстрапирамид- ных нарушений, свойственных истинным нейролептикам.
Серотонин играет существенную роль и в ноцицепции (проведении и восприятии болевых сигналов). Экспериментально показано, что электрической стимуляцией нижней группы ядер шва можно вызвать сильную анальгезию.
Имеются данные о том, что серотонинергическая система мозга принимает участие и в когнитивных процессах. Например, при болезни Альцгеймера наряду с поражением холинергических нейронов происходит частичная дегенерация серотонинергических нейронов.
Источник: Белова Е. И., «Основы нейрофармакологии» 2006