Энергетическое звено метаболизма
Источником энергии, используемым организмом для выполнения всех видов работ является энергия химической связи. Высвобождение
энергии осуществляется в результате окислительно-восстановительного распада простых метаболитов: глюкозы, аминокислот, глицерина, жирных кислот, которые получаются при превращении сложных вещества в пищеварительном тракте.
На I этапе полисахариды расщепляются до моносахаридов (обычно гексоз). Жиры распадаются на глицерин и высшие жирные кислоты, а белки - на составляющие их свободные аминокислоты. Эти процессы в основном являются гидролитическими, и освобождающаяся в небольшом количестве энергия используется в качестве тепла.
На II этапе мономерные молекулы (гексозы, глицерин, жирные кислоты и аминокислоты) подвергаются дальнейшему распаду, в процессе которого образуются богатые энергией фосфатные соединения и ацетил- КоА. В частности, при гликолизе гексозы расщепляются до пировиноград- ной кислоты и далее до ацетил-КоА. Этот процесс сопровождается образованием ограниченного числа богатых энергией фосфатных связей путем субстратного фосфорилирования. Высшие жирные кислоты на этом этапе распадаются до ацетил-КоА, в то время как глицерин окисляется по глико- литическому пути до пировиноградной кислоты и далее до ацетил-КоА.
Использования аминокислот как источника энергии (при дефиците углеводов) осуществляется по разному. Одни аминокислоты непосредственно превращаются в метаболиты цикла Кребса (глутамат, аспартат), другие - опосредованно через глутамат (пролин, гистидин, аргинин), третьи - в пируват и далее в ацетил-КоА (аланин, серин, глицин, цистеин). Ряд аминокислот, в частности лейцин, изолейцин, расщепляется до ацетил- КоА, а из фенилаланина и тирозина, помимо ацетил-КоА, образуется окса- лоацетат через фумаровую кислоту.
Таким образом II этап - это этап образования ацетил-КоА, являющегося по существу единым (общим) промежуточным продуктом катаболизма основных пищевых веществ в клетках.
На III этапе ацетил-КоА подвергаются окислению («сгоранию») в цикле трикарбоновых кислот. Окисление сопровождается образованием восстановленных форм НАДН и ФАДН2. По существу первые три этапа можно определить как процесс катаболического превращения крупных молекул (рис. 43).
На IV этапе осуществляется перенос электронов от восстановленных нуклеотидов на кислород (через дыхательную цепь). Он сопровождается образованием конечного продукта - молекулы воды. Этот транспорт электронов сопряжен с синтезом АТФ в процессе окислительного фосфорили- рования.
Окислительное фосфорилирование самый эффективный способ синтеза АТФ, в результате которого компоненты дыхательной цепи катализируют перенос электронов от НАДН (или восстановленного убихинона) на молекулярный кислород. При этом образуется энергия для синтеза АТФ.
Это постоянно действующий и наиболее эффективный путь энергообразования в клетках всех типов, так как в нем наряду с глюкозой, могут быть использованы не только жирные кислоты, но и кетоновые тела. Подчеркнем, что при снижении парциального давления кислорода до 90 мм.рт.ст. скорость аэробного гликолиза и окислительного фосфорилирования существенно снижаются. Клиническим эквивалентом этого снижения являются слабость, разбитость, плохое самочувствие в целом.
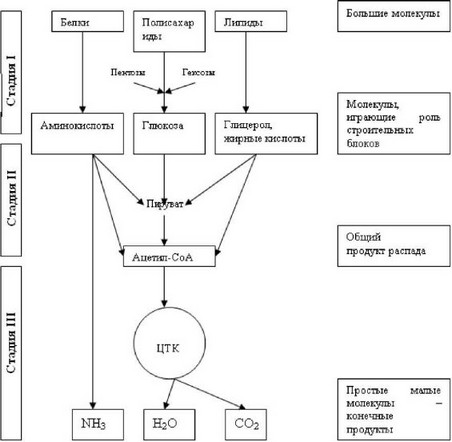
Рис. 43. Этапы катаболического превращения крупных молекул.
Примечание: ЦТК - цикл трикарбоновых веществ.
Помимо основного источника энергии описанного выше существуют альтернативные источники получения энергии:
- Анаэробный гликолиз - при отсутствии или недостатке в клетке кислорода пировиноградная кислота подвергается восстановлению до молочной кислоты (боли в мышцах, возникающие через некоторое время по
сле непривычной интенсивной физической нагрузки, связаны именно с накоплением в них молочной кислоты). Образование молочной кислоты не является конечным продуктом обмена веществ. Под действием лак- татдегидрогеназы молочная кислота окисляется снова в пируват. Кроме того, током крови молочная кислота переносится в печень, где превращается в глюкозу, которая через кровь разносится по всему организму (цикл Кори) (рис. 44). Без существенных последствий для организма анаэробный гликолиз может покрывать кратковременные энергетические нагрузки, даже субмаксимальные. Однако при заболеваниях анаэробный гликолиз не обеспечивает в полной мере потребности клеток в энергии, при этом накопливается молочная кислота, и в результате этого возникает недостаточность функциональных систем, в том числе, не связанных напрямую с пораженной системой или органом. - Субстратное фосфорилирование - образование АТФ в ходе метаболического цикла (переход сукцинат-КоА в сукцинат в цикле Кребса и образование пирувата при гликолизе) (рис. 45). Эти реакции способны на некоторое время поддержать жизнедеятельность организма в отсутствии окислительного фосфорилирования.
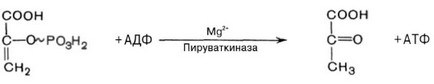
Рис. 45. Пример реакции субстратного фосфорилирования. Образование пирувата в гликолизе.
- Пентозофосфатный путь окисления глюкозы (или «пентозный шунт») - необходим для ресинтеза жирных кислот и предшественников нуклеотидов. При этом образуется НАДФН и продукты способные включаться в гликолиз и далее в цикл трикарбоновых кислот (рис. 46).5—
- Гидролиз креатинфосфата - быстрый и кратковременный путь получения энергии за счет гидролиза креатинфосфата (рис. 47).
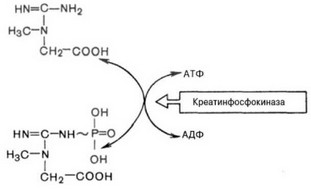
Рис. 47. Ферментативная реакция с участием креатинфосфокиназы.
- Образование инозинмонофосфата в результате конверсии АДФ в АТФ и АМФ (рис. 48).
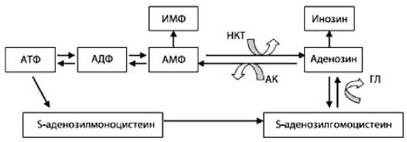
Рис. 48. Основные пути внутриклеточной биотрансформации аденозина-
зина.
Примечание: ИМФ - инозинмонофосфат, АТФ - аденозинтрифосфат, АДФ - аденозиндифосфат, АМФ - аденозинмонофосфат, НКТ - 5-нуклеотидаза, АК - адено- зинкиназа, ГЛ - гидролаза.
- Р-Окисление жирных кислот - происходит в митохондриях, при низкой концентрации пирувата и высоком содержание НАД+ (рис. 49).
Таким образом, основным источником энергии является цикл Кребса сопряженный с окислительным фосфорилировнаием. Главным и быстро мо
билизуемым исходным субстратом служит глюкоза. Ее метаболизм покрывает основной обмен и обеспечивает жизнедеятельность организма. Главным регуляторным механизмом цикла трикарбоновых кислот и, отчасти окислительного фосфорилирования, является кругооброт окислительновосстановительных эквивалентов, которые обозначают отношением НАДН/НАД+.
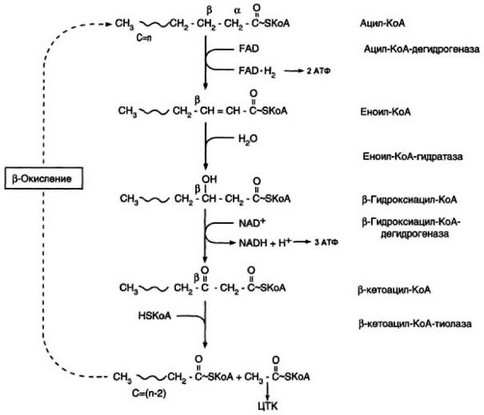
Рис. 49. Р-Окисление жирных кислот (по Березову Т.Т., Коровкину Б.Ф., 1998).
А так же в разделе «Энергетическое звено метаболизма »
- Особенности метаболизма клетки в норме и при патологии
- Пластическое звено метаболизма
- Утилизации продуктов метаболизма
- Метаболические механизмы реактивности клеток иммунной системыИзменение метаболизма в лимфоцитах в процессе их функционирования в норме и при патологии
- Метаболизм гранулоцитов и макрофагов в состоянии относительного покоя и при фагоцитозе
- Значение оксидоредуктаз в системе внутриклеточного метаболизма
- 2.4. Роль витаминов в системе клеточного метаболизма Межвитаминные взаимоотношения
- Глава 3 ВИТАМИНЫ И НАРУШЕНИЯ ФУНКЦИИ ИММУННОЙ СИСТЕМЫ
- Витаминная недостаточность и их клинические проявления
- Клиническая картина гиповитаминозов Гиповитаминоз A
- Гиповитаминоз В1
- Гиповитаминоз В2
- Гиповитаминоз В6
- Г иповитаминоз фолиевой кислоты (В9)
- Гиповитаминоз В12
- Гиповитаминоз C
- Гиповитаминоз D
- Гиповитаминоз E
- Недостаточность карнитина
- Лабораторные методы исследования витаминов Определение аскорбиновой кислоты в моче по Тильмансу
- Определение рибофлавина (витамина В2) в крови по Берчугу, Бессею и Лоури
- Определение рибофлавина (витамина В2) в моче по Е.М. Масленниковой и Л.Г. Г воздовой
- Определение 4-пиридоксиловой кислоты в моче по Хуффу и Перлцвейгу
- 3.4.8. Определение токоферола (витамина Е) в крови в модификации Фридемана