
Молекулярной основой наследственности у всех прокариот и эукариот является особый класс биоорга- нических веществ - нуклеиновые кислоты, подразделяющиеся по своему химическому составу и биологической роли на дезоксирибонуклеиновые кислоты (ДНК) и рибонуклеиновые кислоты (РНК).
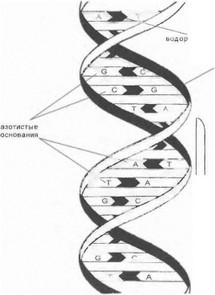
Оба типа нуклеиновых кислот представляют собой нитевидные молекулы, состоящие из отдельных структурных единиц - нуклеотидов, соединенных в многозвеньевую полинуклестидную цепь. Каждый нуклеотид состоит из следующих трех химически различных частей: I) остатков 5-углеродного сахара-дезоксирибо- зы (в ДНК) и рибозы (в РНК), образующих «остов» по- линуклеотидной нити; 2) четырех азотистых оснований- аденина (А), гуанина (G), цитозина (С) и тимина (Т) (в молекуле РНК последнее основание заменено на урацил U), причем каждое азотистое основание ковалентно соединено с первым атомом углерода сахара посредством гликозидной связи; 3) фосфатной группы, соединяющей соседние нуклеотиды в единую цепь посред- ст вом формирования фосфодиэфирных связей между 5’- iTOMOM углерода одного сахара и 3 ’-атомом углерода другого. Первичная структура полинуклеотидных цепей ДНК и РНК представлена на рис. 1.
Запись генетической информации осуществляется линейно от 5’-конца к 3’-концу молекулы нуклеиновой кислоты. В состав одной такой молекулы может входить до многих миллионов нуклеотидов.
В клетке молекулы ДНК существуют в виде спи- рализованной двойной цепи (двойной спирали), нити которой антипараллельны, т.е. имеют противоположную ориентацию. Двойная цепь ДНК образуется благодаря слабым водородным связям между комплементарными основаниями: аденин строго комплементарен тимину, а цитозин - гуанину (рис. 2).
5’ 3’
При определенных условиях указанные водородные связи могут разрываться, приводя к появлению одноцепочечных молекул (денатурация ДНК), а в дальнейшем образовываться вновь между теми же комплементарными участками (ренатурация, или гибридизация ДНК). В процессе гибридизации происходит точное восстановление исходной двойной спирали Д1IK. Именно наличие комплементарное™ обеспечивает как точность самовоспроизводства Д] IK в каждом цикле клеточного деления (этот процесс носит название репликация), так и восстановление нарушенного нуклеотидного состава молекулы ДНК. В связи с комплементарностью нуклеотидов в составе двойной спирали длину молекулы ДНК принято выражать в парах оснований (п.о.), а 1 акже тысячах пар оснований (килобазы, кб) и миллионах пар оснований (мегабазы, мб). В состав ДНК человека как биологического вида входит около 3 миллиардов п.о.
Направленный синтез молекулы ДНК в клетке осуществляется особым ферментом ¦ ДНК-полимеразой. Этот процесс предполагает «расплетение» двойной спирали на участке синтеза и образование особой белковонуклеиновой структуры - репликационной вилки; постепенное продвижение репликационной вилки вдоль двойной спирали сопровождается последовательным присоединением к вновь образуемой цепи оснований, комплементарных однонитевой ДНК-матрице (синтез растущей цепи ДНК всегда протекает строго в направлении от 5’ к 3’). Комплементарный синтез ДНК требует присутствия в среде отдельных «кирпичиков» для удлинения растущей молекулы - четырех видов молекул дезок- сирибонуклеотид-трифосфатов (dATP, dTTP, dCTP и dGTP). Весь процесс инициируется особыми затравками - праймерами, представляющими собой короткие олигонуклеотидные молекулы, комплементарные определенному стартовому участку ДНК-матрицы.
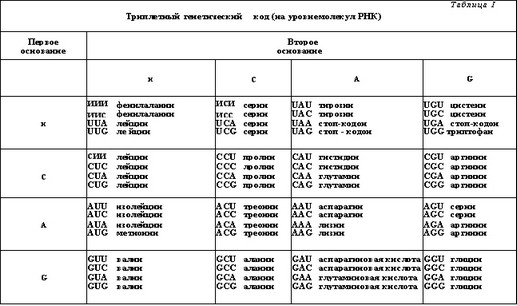
Дискретной единицей наследственности у высших организмов является ген. Совокупность всех генов определенного биологического вида определяется термином геном (иногда данный термин относится к полной I енетической системе отдельной клетки или конкретного организма). Ген в своем наиболее практическом понимании представляет собой строго определенный участок молекулы ДНК, последовательность которого зак- мючает в себе всю информацию, необходимую для синтеза молекулы белка или РНК. Генетическая информа- 11,ия зашифрована посредством универсального для всех живых организмов генетического кода, представляющего собой набор нуклеотидных триплетов - кодонов. Каждый такой триплет каждая последовательность из 3 нуклеотидов) кодирует синтез одной, строго определенной аминокислоты в составе белка (таблица 1).
Считывание кодонов в процессе передачи генетической информации происходит последовательно (принцип линейности генетического кода), и любой нук- псотид может входить в состав только одного кодона (принцип неперекрываемости генетического кода). Генетический код является вырожденным, т.е. допускает кодирование каждой из 20 аминокислот несколькими возможными комбинациями триплетов (всего таких ком- оинаций может быть 64). Расшифровка точной последовательности нуклеотидов определенного информационного участка гена позволяет однозначно идентифицировать последовательность аминокислот в составе соответ- с I дующего полипептидного участка белка и его размер. 11олный гаплоидный геном человека (т.е. кодируемый i gt;, и юй смысловой нитью ДНК) включает, ориентировочно, около 30 000—40 000 генов.
Гены человека и других высших организмов имеют чрезвычайно сложную структурно-функциональную организацию и содержат различные по своей биологической роли нуклеотидные участки. Одни из них (экзо- ны) являются относительно короткими, представляют собой кодирующие последовательности и определяют аминокислотный состав белков; другие участки гена (ин- троны) являются обычно значительно более протяженными и не несут непосредственной информационной нагрузки. Окончательная роль интронов до настоящего времени не установлена; предполагается, что они могут иметь отношение к регуляции экспрессии генов и контролю тонких механизмов «считывания» генетической информации. В состав генов входят также особые регуляторные участки (промоторы, энхансеры, различные сигнальные последовательности), обеспечивающие инициацию, интенсивность и определенную временную последовательность процессов нуклеотидного синтеза на ДНК-матрице, а также модификацию промежуточных полинуклеотидных продуктов. По ориентировочным оценкам, собственно кодирующие последовательности ДНК составляют не более 3-10% всего генома человека.
В любой клетке организма содержится полный набор генов, однако лишь небольшая их часть является функционально активной в каждой конкретной ткани, т.е. экспрессируется. Под экспрессией гена понимают реализацию записанной в нем генетической информации, приводящую к синтезу первичных молекулярных продуктов гена - РПК и белка. Именно временная и тканевая избирательность экспрессии генов определяет специфику дифференцировки и функционирования различных органов, тканей и клеток организма в онтогенезе.
Процесс передачи генетической информации определяется так называемой центральной догмой молекулярной биологии: ДНК^-РНК^-белок. Согласно совре-
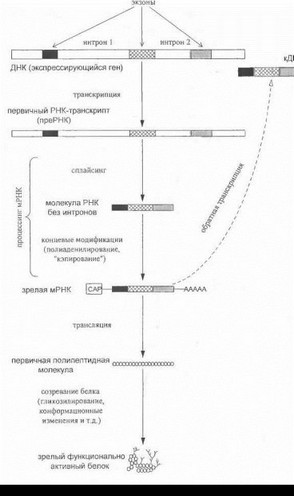
менным представлениям, путь от гена к белку является весьма сложным и распадается не несколько самостоятельных этапов, схематично показанных на рис. 3.
На первом этапе происходит «переписывание» нуклеотидной последовательности гена путем синтеза комплементарной ему молекулы РНК (транскрипция). Транскрипция направляется ферментом РНК-полимера- ;ой и ведет к образованию в ядре клетки молекул первичного РНК-транскрипта (преРНК). Молекула преРНК [ федставляет собой точный слепок ДНК-матрицы транскрибируемого гена. Синтезированная преРНК проходит стадию созревания (процессинг): эта стадия включает в себя как модификацию концевых участков цени (способствующую стабилизации молекулы), так и удаление из первичного РНК-транскрипта некодирующих участков- нитронов.
Процесс «вырезания» интронов, который носит название сплайсинг, является важнейшим звеном созревания преРНК и приводит к тому, что в составе РНК остаются лишь последовательно «сшитые» друг с дру-
- ом смысловые участки, комплементарные экзонам гена. Ключевую сигнальную роль в осуществлении сплайсинга
- грают определенные нуклеотидные последовательности, фланкирующие каждый из экзонов (так называемые сайты сплайсинга); при локализации мутаций в сайтах сплайсинга может происходить нарушение интимных механизмов удаления интронов из состава преРНК и как результат - синтез аномального по структуре пептида. ()бразующаяся после вырезания интронов зрелая РНК носит название информационной, или матричной (мРНК); по своей длине мРНК во много раз короче самого транскрибируемого гена и его первичного РНК- гранскрипта.
Следующий этап передачи генетической информации происходит в цитоплазме. Он заключается в сборке на рибосомах молекул белка по матрице мРНК (процесс трансляции). Аминокислоты транспортируются к рибосомам особым классом молекул - транспортными РНК (тРНК). Каждая тРНК отвечает за транспортировку строго определенной аминокислоты, причем эта специфичность определяется наличием в составе тРНК уникальной 3-нуклеотидной последовательности, называемой антикодоном. По мере продвижения рибосомы вдоль молекулы мРНК антикодоны различных тРНК, несущих «свою» аминокислоту, последовательно распознаются комплементарными им кодонами мРНК. В результате этого происходит последовательное присоединение «нужных» аминокислот к растущей полипептид- ной цепи. Процесс трансляции инициируется триплетом AUG, кодирующим аминокислоту метионин. Таким образом, метиониновый кодон в составе РНК открывает рамку считывания генетической информации; как было указано, это считывание происходит в соответствии с правилом «один триплет - одна аминокислота». Сигналом окончания трансляции служит один из трех особых кодонов (DAA, UAG или UGA), получивших название стоп-кодоны (нонсенс-кодоны); распознавание стоп-ко дона на рибосоме прекращает синтез полипептидной цепи.
По окончании трансляции первичная полипептид- ная молекула претерпевает определенные посттрансляционные модификации, превращаясь в функционально зрелый продукт. «Дозревание» белка происходит, как правило, в соответствующих ррганеллах клетки.
Существует принципиальная возможность другого пути передачи генетической информации (т.е. в направлении, обратном положениям центральной догмы) - от молекулы РНК к молекуле ДНК. В естественных условиях такой путь свойственен некоторым РНК-содер- жащим вирусам, в то время как у высших организмов обратная ч ранскрипция мРНК в ДНК возможна in vitro с участием особого фермента-об/?ш?шой транскриптазы. В результате образуется искусственная молекула ДНК, не содержащая нитронов и полностью комплементарная мРНК-матрице (так называемая комплементарная ДНК, или кДНК) (рис. 3, пунктирная стрелка). Поскольку молекула кДНК представляет собой, образно говоря, концентрированный кодирующий продукт исходного гена, она является весьма ценным материалом при проведении различных молекулярно-генетических исследований (в том числе с целью генодиагностики и 1 енотерапии).
ДНК человека в основной своей части упакована в ядрах клеток в форме компактных структур - хромосом. Каждая хромосома представляет собой одну гигантскую гиперспирализованную молекулу ДНК, образующую сложный комплекс с особыми ядерными белками - гистонами. Во всех ядерных соматических клетках набор хромосом парный (по одному от каждого родителя), в половых - одинарный. В норме у человека 46 хромосом представлены 23 парами, из которых 22 являются иеполовыми (аутосомы) и 1 пара - половые хромосомы (X и Y). Набор половых хромосом детерминируют пол индивидуума (XY - мужской, XX - женский).
В составе каждой хромосомы можно выделить длинное и короткое плечо, разделяемые центральной перетяжкой - центромерой. Хромосомы не идентичны по своим размерам и строению (соотношение длинного и короткого плеча). При использовании специальной дифференциальной окраски хромосомы приобретают поперечно-исчерченный вид, который является их «индивидуальным портретом», позволяющим идентифици- 1 ювать каждую из хромосом при световой микроскопии. Данная картина хромосом определяется их нуклеотидным составом, а также соотношением эухроматина и гетерохроматина (эухроматин - нуклеопротеидный комплекс с низкой плотностью упаковки, содержащий основное число активно транскрибируемых генов; гетерохроматин - конденсированная часть нуклеопротеиндно- го комплекса, содержит в основном некодирующие высокоповторяющиеся последовательности ДНК). В качестве примера на рис. 4 представлена цитогенетическая карта 2-й хромосомы (эта хромосома является одной из наиболее «значимых» применительно к нейрогенетичес- ким заболеваниям). В соответствии с действующей международной цитогенетической номенклагурой (ISCN, 1978) каждый участок хромосомы обозначается символом, отражающим плечо хромосомы (р - короткое, q - длинное), номер сегмента по направлению от центромеры к концевому участку хромосомы (теломере), далее внутри каждого сегмента - номер полосы, а также иногда- номер более мелких субъединиц в составе полос. Так, символ 2р 13.1 подразумевает участок, расположенный в 1-й субъединице 3-й полосы 1-го сегмента короткого плеча хромосомы 2. Центромерные и наиболее терминальные теломерные участки каждого плеча хромосомы обозначаются, соответственно, символами сеп и ter. Например, обозначение 17pter указывает на наиболее дистальный терминальный участок короткого плеча 17-й хромосомы.
Деление соматической клетки и ее ядра (митоз) сопровождается сложными многофазными трансформациями хромосом (рис. 5): 1) в процессе митоза происходит удвоение каждой хромосомы на основе комплементарной репликации молекулы ДНК с образованием двух сестринских нитевидных копий (:хроматид), соединенных в области центромеры; 2) в последующем сестринские хроматиды разъединяются и эквивалентно распределяются по ядрам дочерних клеток.
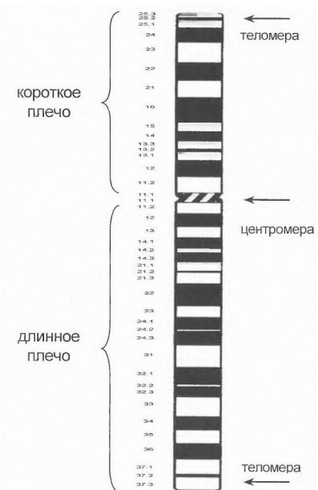
1’ис. 4. Цитогенетическая карта хромосомы 2
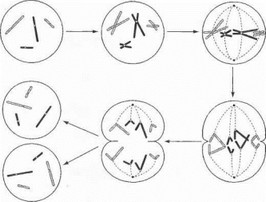
Рис. 5. Схема митоза
Механизм эквивалентного расхождения генетического материала по дочерним клеткам показан на примере 2 пар хромосом
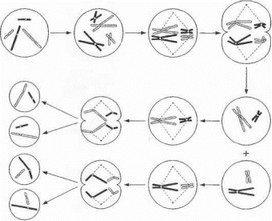
Рис. 6. Схема мейоза
Механизм образования четырёх гаплоидных клеток показан на примере 2 пар хромосом.
В результате в делящихся соматических клетках поддерживается идентичность хромосомного набора и | ппетического материала. Отдельно следует сказать о нейронах - высокодифференцированных постмитотичес- к и \ клетках, не претерпевающих клеточных делений на 111 нтгяжении жизни. Компенсаторные возможности нейронов в ответ на действие повреждающих факторов ог- р: 111 ичиваются внутриклеточной регенерацией и репарацией ДНК в недедящемся ядре, чем в значительной степени обусловлена специфика нейропатологических про- | icccob наследственной и ненаследственной природы.
Совершенно иной тип деления - мейоз - харак-
- срен для половых клеток. Главной особенностью мейоза являются два последовательных деления клетки-
- редшественника и ее ядра, в то время как хромосомы упаиваются лишь однажды. Схематично механизм мей- о::а выглядит следующим образом (рис. 6): 1) в первом делении мейоза дочерние клетки получают из каждой хромосомной пары по одной гомологичной хромосоме, (ч стоящей из удвоенных сестринских хроматид (поскольку при этом число хромосом в дочерних клетках умень- ш; ются вдвое, данное деление является редукционным); ’) но втором делении сестринские хроматиды разъеди- ы я ются и эквивалентно расходятся по образующимся зрелым половым клеткам - гаметам. В результате число х | к iMocoM в гаметах оказывается вдвое меньшим по сравнению с исходной родительской клеткой. После слияния ядер половых клеток при онлодотворении зигота получает стандартный двойной набор хромосом. Данный механизм обеспечивает постоянство числа хромосом у разных поколений организмов, размножающихся половым путем.
Важнейшей биологической ролью мейоза является обеспечение генетического разнообразия особей в результате «перемешивания» отцовских и материнских генов в гамете. Это достигается двумя путями. Во-первых, как показано на рис. 6, в первом делении мейоза распределение отцовских и материнских хромосом по дочерним клеткам происходит случайным образом, в результате чего гаметы несут различные комбинации родительских хромосом. Второй фундаментальный механизм поддержания генетического разнообразия заслуживает того, чтобы быть разобранным более подробно, поскольку он имеет прямое отношение к теме настоящей монографии - ДНК-диагностике. Данный механизм схематично показан на рис. 7.
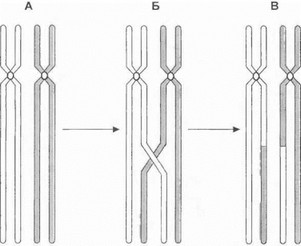
Рис. 7. Кроссинговер между гомологичными хромосомами в мейозе
А. Спаривание гомологичных хромосом. Б. Образование хиазмы и обмен гомологичными участками хромосом. В. Образующиеся в результате кроссинговера рекомбинантные хромосомы.
1} начальной фазе первого деления мейоза гомологичные хромосомы располагаются друг напротив друга и одариваются, образуя одну или несколько зон контакта (хиазм) между отдельными несестринскими хроматида- ми (рис. 7А, Б). Далее пара хроматид, образовавшая хиазму, обменивается участками ДНК - процесс, носящий, название кроссинговер. В результате кроссинговера образуются рекомбинантные хромосомы, состоящие из участков, имеющих происхождение от разных родительских линий (рис.7В). По завершении мейоза рекомбинан- 1 ные хромосомы разойдутся по разным гаметам. Таким образом, кроссинговер представляет собой частный случай генетической рекомбинации - процесса перераспре- 1еления генетического материала родителей при передаче потомству. Важным следствием кроссинговера становится создание новой комбинации генов у потомков при соединении родительских гамет. Поскольку при рекомбинации происходит обмен генетического материала между отцовской и материнской хромосомами, этот феномен всегда должен приниматься во внимание при анализе наследования хромосом в процессе проведения косвенной ДНК-диагностики и расчете генетического сцепления (см. далее).
В клетке имеется около 5% ДНК, не входящей в состав хромосом ядра и локализованной в митохондриях. Митохондриальная ДНК (мтДНК) представляет собой короткую двуцепочечную кольцевую молекулу, которая состоит из 16 569 п.о. и содержит 37 генов, кодирующих синтез некоторых видов рибосомальной и транспортной РНК, а также небольшого числа полипептидов дыхательной цепи митохондрий. Особенностями мтДНК являются: 1) отсутствие некодирующих областей - нитронов; 2) некоторые отличия генетического кода по сравнению с ядерной ДНК; 3) более простая организация и отсутствие связи с белками-гистонами; 4) несовершенство системы репарации ДНК. Два последних фактора лежат в основе того, что повреждаемость (темп мутирования) мтДНК на порядок выше, чем ядерной ДНК. Одна митохондрия (а их число в клетке может достигать многих десятков и сотен) обычно содержит несколько копий мтДНК - в среднем около 5. Поэтому популяция молекул мтДНК в любой клетке и ткани является весьма значительной. Каждая молекула мтДНК реплицируется самостоятельно, и при делении клетки различные молекулы мтДНК вместе с митохондриями в случайном порядке переходят в цитоплазму дочерних клеток. Таким образом, клетки и ткани могут содержать как мтДНК одного вида (гомоплазмия), так и комбинацию нормальной и мутантной мтДНК в различных соотношениях (зе- тероплазмия). Набор митохондрий в зиготе и, следовательно, во всех клетках организма имеет исключительно материнское происхождение - из цитоплазмы яйцеклетки, поэтому мтДНК всегда наследуется по материнской линии. Мутации мтДНК лежат в основе особого класса заболеваний - митохондриальных цитопатий (см. главу 3, раздел 3.8).