Изменение трансмембранного потенциала
Для возбудимых тканей характерно изменение трансмембранного потенциала под действием раздражителей. В том случае, если трансмембранный потенциал увеличивается по отношению к МПП, говорят о гиперполяризации, если уменьшается — о деполяризации, если после деполяризации трансмембранный потенциал вновь увеличивается до МПП — это реполяризация.
Мембрана нейрона способна регулировать два принципиально различающихся по свойствам типа электрических процесса — это локальный ответ (препотенциал или в случае с рецепторами —рецепторный потенциал) и потенциал действия.
Локальный ответ — это потенциал, развивающийся при действии подпороговых, для данной клетки или группы клеток, по силе раздражителей [13, 26,31] (рис. 3.8).
Механизм генерации локального ответа, представленного на рис. 3.8, заключается в следующем.
На начальном этапе осуществляется физическая деполяризация участка мембраны (уменьшается трансмембранный потенциал). Причиной физической деполяризации может служить, например, изменение трансмембранного потенциала соседней клетки. Снижение трансмембранного потенциала вызывает изменение трехмерной структуры белковой молекулы канала. При этом происходит открытие Nav потенциал-зависимых каналов (управляющий фактор — траисмемб- ранный потенциал) и, как следствие, формируется входящий в клетку (по градиенту концентрации) натриевый ток. Натрий «приносит» в клетку положительный заряд и, как следствие, уменьшается трансмембранный потенциал. Таким образом, формируется фаза деполяризации. В дальнейшем Na+ потенциал-зависимые каналы закрываются и тем самым входящий в клетку натриевый ток прекращается. Фаза реполяризации (следующая фаза после деполяризации) формируется за счет открытия К+-потенциал, зависимых каналов, при этом выходящий ток К* «выносит» положительный заряд. При приближении значения трансмембранного потенциала к значению МПП калиевые каналы закрываются и калиевый ток из клетки уменьшается. Благодаря описанным выше механизмам формируется двухфазный (деполяризация и реполяризация) локальный ответ.
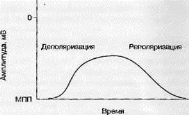
Рис. 3.8. Локальный ответ нейрона
Локальные ответы подчиняются закону силы, что служит предпосылкой для одного из вариантов кодирования информации в нервной системе [13].
I
Закон силы: чем больше сила раздражителя, тем больше величина ответной реакции.
Следовательно, в амплитуде локального ответа закодирована сила раздражителя.
В соответствии с законом силы функционируют сложные структуры, например скелетная мышца. Амплитуда ее сокращений от минимальных (пороговых) величин постепенно увеличивается с увеличением силы раздражителя де субмаксимальных и максимальных значений. Это объясняется тем, что скелетная мышца состоит из множества мышечных волокон, каждое из которых имеет свою возбудимость. Поэтому вначале, когда сила раздражителя пороговая, отвечают только те мышечные волокна, которые имеют самую высокую возбудимость. С увеличением силы раздражителя в реакцию вовлекается все большее и большее количество мышечных волокон, поскольку сила раздражителя для них становится пороговой или сверхпороговой. Когда в реакцию вовлечены все мышечные волокна (для всех раздражитель становится пороговым или сверхпороговым), составляющие данную мышцу, дальнейшее увеличение силы раздражителя не приводит к увеличению амплитуды сокращения [27,32].
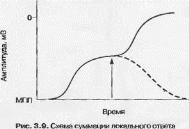
Другим важным свойством локального ответа является его способность к суммации [14,26,32] (рис. 3.9).
На представленном рис. 3.9 видно, что после дополнительного деполяризующего воздействия, время нанесение которого отмечено стрелкой, дальнейшая деполяризация начинается не с уровня МПП, а с уровня, уже достигнутого предыдущим воздействием локального ответа (пунктирной линией отмечено изменение трансмембранного потенциала при отсутствии дополнителного раздражителя).
Амнлитуда локального ответа не может расти до бесконечности. На определенном этапе значения трансмембранного потенциала достигают критическпгоуровня деполяризации (КУД) и процесс деполяризации становится самоусиливаюшимся, что свидетельствует о появлении качественно нового электрического процесса на мембране — потенциала действия (рис. 3.10). КУД - это своего рода граница, порог, разделяющий два процесса, протекающих на мембране одной клетки. Таким образом, тот раздражитель, который деполяризует мембрану до уровня КУД, и будет являться пороговым. А соответственно, разность потенциалов между МПП и КУД — пороговым потенциалом, который характеризует возбудимость клетки.
На первом этапе локальный ответ представляет собой деполяризацию, при этом появляется входящий натриевый ток. Важно отметить, что локальный ответ возникает не только при подпороговом, но и при пороговом и сверхпороговом раздражении и является неотъемлемой частью потенциала действия.
При достижении КУД деполяризация принимает самовоспроизво- дящийся характер [14, 27, 32]. В этот момент натрий «лавинообораз- но» поступает в клетку, что связано с массовым открытием натриевых потенциал-зависимых каналов. Открытие каналов вызывает деполяризацию, а деполяризация мембраны вызывает открытие натриевых потенциал-зависимых каналов. На рис. 3.10 видно быстрое уменыие- ние мембранного потенциала и даже перезарядку мембраны: внутренняя ее часть на некоторое время становится заряженной положительно, а внешняя — отрицательно (овершут). Б отличие от локального ответа скорость и величина деполяризации не зависят от силы раздражителя, т. е. работает закон «все или ничего».
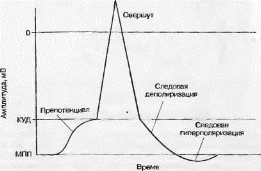
Рис. 3.10. Схема потенциала действия ответа
За фазой деполяризации наступает фаза реполяризации. Фаза реполяризации характеризуется восстановлением мембранного потенциала под действием выходящего из клетки калиевого тока. В механизме реполяризации участвуют не только открывающиеся потенциал-за- висимые каналы, через которые ионы калия покидают клетку, но и натриевые потенциал-зависимые каналы, которые закрываются (ин- кативируются), что обеспечивает замедление деполяризации вплоть до ее полной остановки. После увеличения в ходе реноляризаиии трансмембранного потенциала больше КУД последовательно формируются два вида следовых потенциалов — следовая деполяризация и следовая гшерполяризация.
Следовая деполяризация является продолжением фазы реполяризации и характеризуется более медленным (по сравнению с фазой ре- поляризации) восстановлением потенциала покоя (выходящий калиевый ток здесь по-прежнему является основным).
Следовая деполяризация переходит в следовую гиперполяризацию, представляющую собой временное увеличение мембранного потенциала выше уровня МПП. В некоторых случаях следовая деполяризация может переходить в следовую гиперполяризацию, затем иногда возникает новая деполяризация, лишь после этого происходит полное восстановление потенциала покоя [26, 27].
В основе изменения трансмембранного потенциала лежат ионные механизмы, которые, в свою очередь, основываются на изменении проницаемости мембраны для ионов, в первую очередь натрия и калия.
При действии на клетку раздражителя проницаемость мембраны для ионов Na* резко повышается за счет активации (открывания) натриевых каналов (хемо- или потенциал-зависимых). При этом ионы Na+ по градиенту концентрации попадают в клетку. Поскольку поток Na+ в клетку начинает превышагь калиевый ток из клетки, то происходит постепенное снижение разности потенциалов между внутри- и внеклеточной средой, приводящее к реверсии — изменению знака мембранного потенциала. Здесь внутренняя поверхность мембраны становится положительной по отношению к ее внешней поверхности. Указанные изменения мембранного потенциала соответствуют восходящей фазе потенциала действия, и в частности овершуту. Мембрана высокопроницаема для ионов NaT лишь очень короткое время: не более 0,5 мс. После этого проницаемость мембраны для ионов Na+ понижается, а для ионов К+ возрастает: трансмембранный потенциал увеличивается. В течение потенциала действия в клетку поступает значительное количество Na+, а ионы К+ покидают клетку. Восстановление клеточного ионного баланса осуществляется благодаря работе Na4. К+-насоса, активность которого возрастает при повышении внутренней концентрации ионов Na+ и увеличении внеклеточной концентрации ионов К\ Благодаря работе Na1, К+-насоса первоначальная концентрация этих ионов во внутриклеточном пространстве постепенно восстанавливается. За один потенциал действия концентрация вне- и внутриклеточных ионов существенно нс меняется и нейрон способен сгенерировать большое количество потенциалов, прежде чем концентрация ионов существенно изменится.
Итогом вышеперечисленных процессов является генерация потенциала действия с возврашением мембраны в исходное состояние.
Важную роль в генерации потенциала действия и в целом ряде элек- трифизиологических феноменов играют натриевые потенциал-зави- симые каналы. Особенностью строения этих каналов является наличие двух ворот, т и к (рис. 3.11).
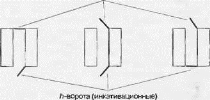
Первые считаются быстрыми, а вторые — медленными. В зависимости от того, какие ворота открыты, а какие закрыты, клетка может менять свое функциональное состояние [13, 26, 27, 32]. Ниже представлены этапы функционирования каналов.
Рис. 3.11. Функционирование натриевого потенциал-зависимого канала
Основные состояния натриевых каналов представлены на рис. 3.10.
- В покое канал закрыт (закрыты m-ворота). Движение ионов через канал невозможно.
- При деполяризации 7Я-ворота (быстрые ворота) открываются и канал активируется (через него возможен ток Na+). Однако //-ворота (медленные ворота) закрываются. За то время, что быстрые ворота уже открыты, а медленные еще не закрылись, канал акти-
- При длительной деполяризации закрываются h-ворота (инактивирующие ворота), расположенные у внутренней стороны мембраны, и канал инактивируется. Вновь открыть его для тока ионов можно лишь сняв инактивацию, что достигается реполяризацией до уровня потенциала покоя. Вновь открываются /z-ворота и закрываются m-ворота. Канал снова потенциально активен [26,32]. Закон «все или ничего»: подпороговые раздражители не вызывают ответной реакции («ничего»), на пороговые раздражители возникает максимальная ответная реакция («все»).
Закон «все или ничего» далеко не абсолютен. Подобные реакции наблюдаются на уровне электрогенеза в клетке, но, к примеру, раздражители подпороговой силы вызывают в ткани изменения мембранного потенциала покоя в виде возникновения местного возбуждения (локального ответа).
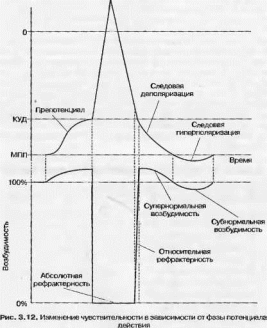
Возбудимость в различные фазы потенциала действия не одинакова и меняется в очень широких пределах (рис. 3.12). Так, во время локального ответа возбудимость повышена. Б этот период клетка может быть возбуждена (т. е. сгенерировать потенциал действия) даже в ответ на субпороговый раздражитель (вспомните суммацию локальных ответов). Когда мембранный потенциал достигает уровня КУД, происходит открытие натриевых каналов, следовательно новый раздражитель не вызовет ответную реакцию, так как каналы уже открыты. В дальнейшем ответная реакция невозможна по другой причине: натриевые каналы хотя и закрыты, но инактивированы (ft-ворота закрыты) и, следовательно, возбудить клетку нельзя. Эта фаза называется фазой абсолютной рефрактерности. При приближении мембранного потенциала к уровню КУД инактивация частично снимается и клетку можно возбудить действуя сверхпороговым раздражителем. Эта фаза получила название относительной рефрактерности. Как только мембранный потенциал превосходит КУД, инактивация снимается, а поскольку разность между КУД и мембранным потенциалом невелика, то клетку можно возбудить субпороговым стимулом. Это фаза супер- нормальной (сверхнсрмальной возбудимости). Когда мембранный потенциал превышает МПП, развивается фаза субгюрмальной возбудимости. Во время этой фазы клетку можно возбудить лишь сверхпороговым раздражителем [32].
Задания и вопросы для проверки усвоения знаний
- Выберите правильный вариант ответа.
- Деполяризацию вызывает:
а) входящий натриевый ток;
б) выходящий натриевый ток;
в) входящий калиевый ток;
г) выходящий калиевый ток;
д) ни один из вышеперечисленных.
- Реполяризацию вызывает:
а) входящий натриевый ток;
б) выходящий натриевый ток;
в) входящий калиевый ток;
г) выходящий калиевый ток;
д) ни один из вышеперечисленных.
- В фазу деполяризации (выберите два ответа):
а) m-ворота открыты;
б) й-ворота открыты,-
в) те-ворота закрыты;
г) й-ворота закрыты;
д) ни одно не верно.
- В фазу абсолютной рефрактерности нейрон можно возбудить:
а) сверхпороговым раздражителем;
б) пороговым раздражителем;
в) подпороговым раздражителем;
г) ни одним из перечисленных выше.
- В фазу относительной рефрактерности нейрон можно возбудить:
а) сверхпороговым раздражителем;
б) пороговым раздражителем;
в) подпороговым раздражителем;
г) ни одним из перечисленных выше.
II. Выполните следующие задания и ответьте на вопросы.
- Приведите классификацию каналов мембраны.
- Перечислите фазы локального ответа.
- Опишите «закон силы».
- Дайте характеристику ионным токам при потенциале действия.
- В чем сущность закона «все или ничего»?
Список тем для подготовки реферативных докладов
- История изучения электрических процессов у живых организмов.
- Электрогенез в нервной ткани.
- Методы исследования потенциала действия.
- Влияние различных химических веществ на генерацию потенциала действия.
- Строение каналов мембраны.
Источник: Щербатых Ю. В., Туровский Я. А., «Физиология центральной нервной системы для психологов» 2007