
Емкость мембраны и работа метаболических ионных насосов приводят к накоплению потенциальной электрической энергии на клеточной мембране в форме потенциала покоя. Эта энергия может освобождаться в виде специфических электрических сигналов (потенциала действия), характерных для возбудимых тканей: нервной, мышечной, некоторых рецепторных и секреторных клеток. Под потенциалом действия понимают быстрое колебание потенциала покоя, сопровождающееся, как правило, перезарядкой мембраны. Форма потенциала действия аксона и терминология, используемая для описания потенциала действия, приведены на рис. 2.10.
 Рис. 2.10. Потенциал действия одиночной клетки и его фазы.
Рис. 2.10. Потенциал действия одиночной клетки и его фазы.
А — реакция клеточной мембраны на деполяризующий стимул; Б — величина деполяризующего тока; 1 — локальный ответ; 2 — быстрая деполяризация; 3 — реверсия; 4 — реполяризация; 5 — положительный следовой потенциал. По оси абсцисс — отметка времени.
Для правильного понимания процессов, происходящих при генерации потенциала действия, используем схему опыта, приведенную на рис. 2.5. Если через стимулирующий электрод подавать короткие толчки гиперполяризующего тока, то можно зарегистрировать увеличение мембранного потенциала, пропорциональное амплитуде подаваемого тока; при этом мембрана проявляет свои емкостные свойства — замедленное нарастание и снижение мембранного потенциала (см. рис. 2.6).
Ситуация будет изменяться, если через стимулирующий электрод подавать короткие толчки деполяризующего тока. При небольшой (подпороговой) величине деполяризующего тока мембрана ответит пассивной деполяризацией и проявит емкостные свойства. Подпороговое пассивное поведение клеточной мембраны называется элек- тротоническим, или электротоном. Увеличение деполяризующего тока приведет к появлению активной реакции клеточной мембраны в форме повышения натриевой проводимости (gNa+). При этом проводимость клеточной мембраны не будет подчиняться закону Ома. Отклонение от пассивного поведения проявляется обычно при 50—80% значении порогового тока. Активные подпороговые изменения мембранного потенциала называются локальным ответом.
Смещение мембранного потенциала до критического уровня приводит к генерации потенциала действия. Минимальное значение тока, необходимого для достижения критического потенциала, называют пороговым током. Следует подчеркнуть, что не существует абсолютных значений величины порогового тока и критического уровня потенциала, поскольку эти параметры зависят от электрических характеристик мембраны и ионного состава окружающей внешней среды, а также от параметров стимула. Зависимость между величиной стимулирующего тока и временем его действия рассматривается в разделе 2.1.5.
В опытах Ходжкина и Хаксли был обнаружен, на первый взгляд, удивительный эффект. Во время генерации потенциала действия мембранный потенциал уменьшался не просто до нуля, как следовало бы из уравнения Нернста, но изменил свой знак на противоположный.
Анализ ионной природы потенциала действия, проведенный первоначально Ходжкиным, Хаксли и Катцем, позволил установить, что фронт нарастания потенциала действия и перезарядка мембраны (овершут) обусловлены движением ионов натрия внутрь клетки. Как уже указывалось выше, натриевые каналы оказались электроуправ- ляемыми. Деполяризующий толчок тока приводит к активации натриевых каналов и увеличению натриевого тока. Это обеспечивает локальный ответ. Смещение мембранного потенциала до критического уровня приводит к стремительной деполяризации клеточной мембраны и обеспечивает фронт нарастания потенциала действия. Если удалить ион Na+ из внешней среды, то потенциал действия не возникает. Аналогичный эффект удавалось получить при добавлении в перфузи- онный раствор ТТХ (тетродотоксин) — специфического блокатора натриевых каналов (см. табл. 2.1). При использовании метода «voltage- damp» было показано, что в ответ на действие деполяризующего тока
Рис. 2.11. Действие деполяризующего стимула (А), ионные токи (Б) и изменение проводимости клеточной мембраны (В) для ионов Na+ (gNa , INa+) и К+ (gK+, IK*) во время действия деполяризующего стимула.
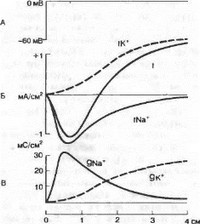 через мембрану протекает кратковременный (1—2 мс) входящий ток, который сменяется через некоторое время выходящим током (рис. 2.11). При замене ионов натрия на другие ионы и вещества, например холин, удалось показать, что входящий ток обеспечивается натриевым током, т. е. в ответ на деполяризующий стимул происходит повышение натриевой проводимости (gNa+). Таким образом, развитие фазы деполяризации потенциала действия обусловлено повышением натриевой проводимости.
через мембрану протекает кратковременный (1—2 мс) входящий ток, который сменяется через некоторое время выходящим током (рис. 2.11). При замене ионов натрия на другие ионы и вещества, например холин, удалось показать, что входящий ток обеспечивается натриевым током, т. е. в ответ на деполяризующий стимул происходит повышение натриевой проводимости (gNa+). Таким образом, развитие фазы деполяризации потенциала действия обусловлено повышением натриевой проводимости.
Критический потенциал определяет уровень максимальной активации натриевых каналов. Если смещение мембранного потенциала достигает значения критического уровня потенциала, то процесс поступления ионов Na+ в клетку лавинообразно нарастает. Система начинает работать по принципу положительной обратной связи, т. е. возникает регенеративная (самоусиливающаяся) деполяризация.
Перезарядка мембраны, или овершут, весьма характерна для большинства возбудимых клеток. Амплитуда овершута характеризует состояние мембраны и зависит от состава вне- и внутриклеточной среды. На высоте овершута потенциал действия приближается к равновесному натриевому потенциалу, поэтому происходит изменение знака заряда на мембране.
Экспериментально было показано, что амплитуда потенциала действия практически не зависит от силы стимула, если он превышает пороговую величину. Поэтому принято говорить, что потенциал действия подчиняется закону "все или ничего”.
На пике потенциала действия проводимость мембраны для ионов натрия (gNa*) начинает быстро снижаться. Этот процесс называется инактивацией. Скорость и степень натриевой инактивации зависят от величины мембранного потенциала, т. е. они потенциалзависимы. При постепенном уменьшении мембранного потенциала до —50 мВ
(например, при дефиците кислорода, действии некоторых лекарственных веществ) система натриевых каналов полностью инактивируется и клетка становится невозбудимой.
Потенциал зависимость активации и инактивации в большой степени обусловлена концентрацией ионов кальция. При повышении концентрации кальция значение порогового потенциала увеличивается, при понижении — уменьшается и приближается к потенциалу покоя. При этом в первом случае возбудимость уменьшается, во втором — увеличивается.
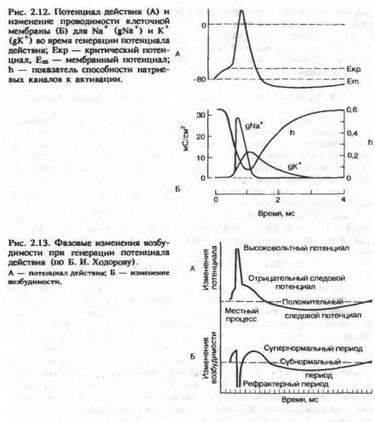
После достижения пика потенциала действия происходит реполяризация, т. е. мембранный потенциал возвращается к контрольному значению в покое. Рассмотрим эти процессы подробнее. Развитие потенциала действия и перезарядка мембраны приводят к тому, что внутриклеточный потенциал становится еще более положительным, чем равновесный калиевый потенциал, и, следовательно, электрические силы, перемещающие ионы калия через мембрану, увеличиваются. Максимума эти силы достигают во время пика потенциала действия. Кроме тока, обусловленного пассивным передвижением ионов калия, был обнаружен задержанный выходящий ток, который также переносился ионами К+, что было показано в опытах с применением изотопа К+. Этот ток достигает максимума спустя 5—8 мс от начала генерации потенциала действия. Введение тетраэтиламмония (ТЭА) — блокатора калиевых каналов —.замедляет процесс реполяризации. В обычных условиях задержанный выходящий калиевый ток существует некоторое время после генерации потенциала действия и это обеспечивает гиперполяризацию клеточной мембраны, т. е. положительный следовой потенциал. Положительный следовой потенциал может возникать и как следствие работы натриево-электрогенного насоса. На рис. 2.12 показано изменение проводимости клеточной мембраны для ионов натрия и калия в различные фазы потенциала действия.
Инактивация натриевой системы в процессе генерации потенциала действия приводит к тому, что клетка в этот период не может быть повторно возбуждена, т. е. наблюдается состояние абсолютной рефракт ерност и.
Постепенное восстановление потенциала покоя в процессе реполяризации дает возможность вызвать повторный потенциал действия, но для этого требуется сверхпороговый стимул, так как клетка находится в состоянии относительной рефрактерности.
Исследование возбудимости клетки во время локального ответа или во время отрицательного следового потенциала показало, что генерация потенциала действия возможна при действии стимула ниже порогового значения. Это состояние супернормальности, или экзальтации. На рис. 2.13 показано изменение возбудимости во время генерации потенциала действия.
Продолжительность периода абсолютной рефрактерности ограничивает максимальную частоту генерации потенциалов действия данным типом клеток. Например, при продолжительности периода абсолютной рефрактерности 4 мс максимальная частота равна 250 Гц.
Н. Е. Введенский ввел понятие лабильности, или функциональной подвижности, возбудимых тканей. Мерой лабильности является количество потенциалов действия, которое способна генерировать возбудимая ткань в единицу времени. Очевидно, что лабильность возбудимой ткани в первую очередь определяется продолжительностью периода рефрактерности. Наиболее лабильными являются волокна слухового нерва, в которых частота генерации потенциалов действия достигает 1000 Гц.
Таким образом, генерация потенциала действия в возбудимых мембранах возникает под влиянием различных факторов и сопровождается повышением проводимости клеточной мембраны для ионов натрия, входом их внутрь клетки, что приводит к деполяризации клеточной мембраны и появлению локального ответа. Этот процесс может достигнуть критического уровня деполяризации, по- еле чего проводимость мембраны для натрия увеличивается до максимума, мембранный потенциал при этом приближается к натриевому равновесному потенциалу. Через несколько миллисекунд происходит инактивация натриевых каналов, активация калиевых каналов, увеличение выходящего калиевого тока, что приводит к реполяризации и восстановлению исходного потенциала покоя.