
В реализации информационно-управляющей функции нервной системы значительная роль принадлежит процессам интеграции и координации деятельности отдельных нервных клеток и нейронных ансамблей, которые основаны на особенностях взаимодействия информационных потоков на уровне нервных клеток и рефлекторных дуг. Конструктивные особенности афферентных, промежуточных (центральных) и эфферентных нейронов обеспечивают широкий диапазон иррадиации и концентрации возбуждения на основе двух кардинальных принципов: дивергенции и конвергенции. Дивергенцией называется способность нервной клетки устанавливать многочисленные синаптические связи с различными нервными клетками. Благодаря этому одна нервная клетка может участвовать в нескольких различных реакциях, передавать возбуждение значительному числу других нейронов, которые могут возбудить большее количество нейронов, обеспечивая широкую иррадиацию возбудительного процесса в центральных нервных образованиях.
Процессы конвергенции заключаются в схождении различных импульсных потоков от нескольких нервных клеток к одному и тому же нейрону (см. раздел 4.1.4). Процесс конвергенции характерен не только для однотипных нервных клеток. Например, на мотонейронах спинного мозга, кроме первичных афферентных волокон, конвергируют волокна различных нисходящих трактов от супраспинальных и собственно спинальных центров, а также от возбуждающих и тормозных вставочных промежуточных нейронов. В результате мотонейроны спинного мозга выполняют функцию общего конечного пути для многочисленных нервных образований, включая и надсегментный аппарат головного мозга, имеющих отношение к регуляции двигательной функции.
В координационной деятельности центральных нервных образований значительная роль взаимодействия рефлексов, которая проявляется в различных эффектах (в облегчении, или суммации, и в угнетении, или подавлении, возбуждения).
Примером координационного взаимодействия рефлексов является реципрокная иннервация мышц-антагонистов. Известно, что сгибание или разгибание конечностей осуществляется благодаря согласованной работе двух функционально антагонистических мышц: сгибателей и разгибателей. Координация обеспечивается организацией антагонистических отношений между мотонейронами сгибателей и разгибателей, иннервирующих соответствующие мышцы. Реципрокные функциональные отношения складываются в сегментарных структурах спинного мозга благодаря включению в дугу спинномозгового рефлекса дополнительного элемента — специального тормозного нейрона (клетка Реншоу). Схематически в общем виде этот механизм выглядит следующим образом: сигнал от афферентного звена через обычную вставочную (промежуточную) нервную клетку вызывает возбуждение мотонейрона, иннервирующего мышцу-сгибатель, а через дополнительную клетку Реншоу тормозит мотонейрон, иннервирующий мышцу-разгибатель. Так происходит координированное сгибание конечности; напротив, при выполнении разгибания конечности возбуждается мотонейрон мышцы-разгибателя, а через вставочную клетку Реншоу тормозится, угнетается мотонейрон мышцы-сгибателя (рис. 4.3). Таким образом акт сгибания конечности, представляющий собой работу мышц-антагонистов, обеспечивается формированием реципрокных антагонистических отношений между нервными центрами соответствующих мышц благодаря наличию специальных тормозных нейронов — клеток Реншоу.
В центральной нервной системе весьма распространены интегративные феномены — посттетаиическая потенциация, окклюзия, облегчение.
Феномен посттетанической потенциации проявляется следующим образом. Раздражая стимулами редкой частоты афферентный нерв, можно получить некоторый рефлекс определенной интенсивности. Если затем этот нерв в течение некоторого времени подвергать высокочастотному ритмическому раздражению (300—400 стимулов в секунду), то повторное редкое ритмическое раздражение приведет к резкому усилению реакции (рис. 4.4).
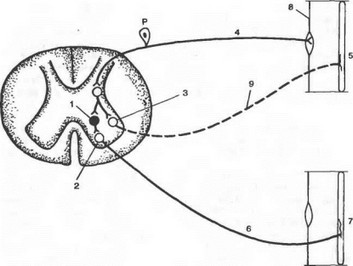
Рис. 4.Э. Реципрокная иннервация мышц-антагонистов.
1 — тормозная клетка Рсншоу; 2 — мотонейрон мышцы-сгибателя; 3 — мотонейрон мышцы-разгибателя; 4 — афферентное волокно от мышцы разгибателя; 5 — мышца-разгибатель; 6 — двигательное (эфферентное) волокно мышцы-сгибателя; 7 — мышца-сгибатель; 8 — мышечный рецептор; 9 — эфферентное нервное волокно мышцы-разгибателя.

Рис. 4.4. Феномен посгтетанической потенциации.
1 — тестовый ответ; 2 — тетаническая стимуляция; 3 — потенцированный ответ нервной клетки.
Если два нервных центра рефлекторных реакций имеют частично перекрываемые рецептивные поля, то при совместном раздражении обоих рецептивных полей реакция будет меньше, чем арифметическая сумма реакций при изолированном раздражении каждого из рецептивных полей — феномен окклюзии (рис. 4.5). В ряде случаев вместо такого ослабления реакции при совместном раздражении рецептивных полей двух рефлексов можно наблюдать феномен облегчения (т. е. суммарная реакция выше суммы реакции при изолированном раздражении этих рецептивных полей). Это результат того, что часть общих для обоих рефлексов нейронов при изолиро-
Рис. 4.5. Феномен окклюзии. РП|.2 — рецептивное поле.
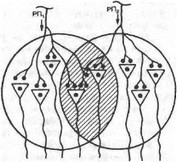 ванном раздражении оказывает подпороговый эффект для вызывания рефлекторных реакций. При совместном раздражении они суммируются и достигают пороговой силы, в результате конечная реакция оказывается больше суммы изолированных реакций.
ванном раздражении оказывает подпороговый эффект для вызывания рефлекторных реакций. При совместном раздражении они суммируются и достигают пороговой силы, в результате конечная реакция оказывается больше суммы изолированных реакций.
Процессы конвергенции заключаются в схождении различных импульсных потоков от нескольких нервных клеток к одному и тому же нейрону (см. раздел 4.1.4). Процесс конвергенции характерен не только для однотипных нервных клеток. Например, на мотонейронах спинного мозга, кроме первичных афферентных волокон, конвергируют волокна различных нисходящих трактов от супраспинальных и собственно спинальных центров, а также от возбуждающих и тормозных вставочных промежуточных нейронов. В результате мотонейроны спинного мозга выполняют функцию общего конечного пути для многочисленных нервных образований, включая и надсегментный аппарат головного мозга, имеющих отношение к регуляции двигательной функции.
В координационной деятельности центральных нервных образований значительная роль взаимодействия рефлексов, которая проявляется в различных эффектах (в облегчении, или суммации, и в угнетении, или подавлении, возбуждения).
Примером координационного взаимодействия рефлексов является реципрокная иннервация мышц-антагонистов. Известно, что сгибание или разгибание конечностей осуществляется благодаря согласованной работе двух функционально антагонистических мышц: сгибателей и разгибателей. Координация обеспечивается организацией антагонистических отношений между мотонейронами сгибателей и разгибателей, иннервирующих соответствующие мышцы. Реципрокные функциональные отношения складываются в сегментарных структурах спинного мозга благодаря включению в дугу спинномозгового рефлекса дополнительного элемента — специального тормозного нейрона (клетка Реншоу). Схематически в общем виде этот механизм выглядит следующим образом: сигнал от афферентного звена через обычную вставочную (промежуточную) нервную клетку вызывает возбуждение мотонейрона, иннервирующего мышцу-сгибатель, а через дополнительную клетку Реншоу тормозит мотонейрон, иннервирующий мышцу-разгибатель. Так происходит координированное сгибание конечности; напротив, при выполнении разгибания конечности возбуждается мотонейрон мышцы-разгибателя, а через вставочную клетку Реншоу тормозится, угнетается мотонейрон мышцы-сгибателя (рис. 4.3). Таким образом акт сгибания конечности, представляющий собой работу мышц-антагонистов, обеспечивается формированием реципрокных антагонистических отношений между нервными центрами соответствующих мышц благодаря наличию специальных тормозных нейронов — клеток Реншоу.
В центральной нервной системе весьма распространены интегративные феномены — посттетаиическая потенциация, окклюзия, облегчение.
Феномен посттетанической потенциации проявляется следующим образом. Раздражая стимулами редкой частоты афферентный нерв, можно получить некоторый рефлекс определенной интенсивности. Если затем этот нерв в течение некоторого времени подвергать высокочастотному ритмическому раздражению (300—400 стимулов в секунду), то повторное редкое ритмическое раздражение приведет к резкому усилению реакции (рис. 4.4).
Рис. 4.Э. Реципрокная иннервация мышц-антагонистов.
1 — тормозная клетка Рсншоу; 2 — мотонейрон мышцы-сгибателя; 3 — мотонейрон мышцы-разгибателя; 4 — афферентное волокно от мышцы разгибателя; 5 — мышца-разгибатель; 6 — двигательное (эфферентное) волокно мышцы-сгибателя; 7 — мышца-сгибатель; 8 — мышечный рецептор; 9 — эфферентное нервное волокно мышцы-разгибателя.
Рис. 4.4. Феномен посгтетанической потенциации.
1 — тестовый ответ; 2 — тетаническая стимуляция; 3 — потенцированный ответ нервной клетки.
Если два нервных центра рефлекторных реакций имеют частично перекрываемые рецептивные поля, то при совместном раздражении обоих рецептивных полей реакция будет меньше, чем арифметическая сумма реакций при изолированном раздражении каждого из рецептивных полей — феномен окклюзии (рис. 4.5). В ряде случаев вместо такого ослабления реакции при совместном раздражении рецептивных полей двух рефлексов можно наблюдать феномен облегчения (т. е. суммарная реакция выше суммы реакции при изолированном раздражении этих рецептивных полей). Это результат того, что часть общих для обоих рефлексов нейронов при изолиро-
Рис. 4.5. Феномен окклюзии. РП|.2 — рецептивное поле.
ванном раздражении оказывает подпороговый эффект для вызывания рефлекторных реакций. При совместном раздражении они суммируются и достигают пороговой силы, в результате конечная реакция оказывается больше суммы изолированных реакций.