Вестибулярная система
Вестибулярная система играет наряду со зрительной и соматосенсорной системами ведущую роль в пространственной ориентировке человека. Она получает, передает и анализирует информацию об ускорениях или замедлениях, возникающих в процессе прямолинейного или вращательного движения, а также при изменении положения головы в пространстве. При равномерном движении или в условиях покоя рецепторы вестибулярной сенсорной системы не возбуждаются. Импульсы от вестибулоре- цепторов вызывают перераспределение тонуса скелетной мускулатуры, что обеспечивает сохранение равновесия тела. Эти влияния осуществляются рефлекторным путем через ряд отделов ЦНС.
Строение и функции рецепторов вестибулярной системы. Периферическим отделом вестибулярной системы является вестибулярный аппарат, расположенный в лабиринте пирамиды височной кости. Он состоит из преддверия (vesti- bulum) и трех полукружных каналов (canales cemicircularis). Кроме вестибулярного аппарата, в лабиринт входит улитка, в которой располагаются слуховые рецепторы. Полукружные каналы (рис. 14.17) располагаются в трех взаимно перпендикулярных плоскостях: верхний — во фронтальной, задний — в сагиттальной и латеральный — в горизонтальной. Один из концов каждого канала расширен (ампула).
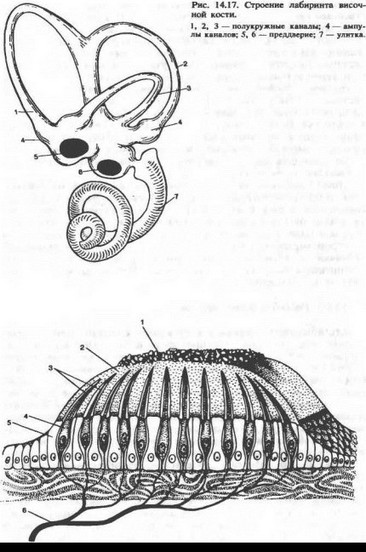
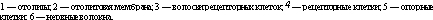
Вестибулярный аппарат включает в себя также два мешочка: сферический (sacculus) и эллиптический, или маточку (utriculus). Первый из них лежит ближе к улитке, а второй — к полукружным каналам. В мешочках преддверия находится отолитовый аппарат: скопления рецепторных клеток (вторично-чувствую- щие механорецепторы) на возвышениях, или пятнах (macula sac- culi, macula utriculi). Выступающая в полость мешочка часть рецепторной клетки оканчивается одним более длинным подвижным волоском и 60—80 склеенными неподвижными волосками. Эти волоски пронизывают желеобразную мембрану, содержащую кристаллики карбоната кальция — отолиты. Возбуждение волоско- вых клеток преддверия происходит вследствие скольжения отоли- товой мембраны по волоскам, т. е. их сгибания (рис. 14.18).
В перепончатых полукружных каналах, заполненных, как и весь лабиринт, плотной эндолимфой (ее вязкость в 2—3 раза больше, чем у воды), рецепторные волосковые клетки сконцентрированы только в ампулах в виде крист (cristae ampularis). Они также снабжены волосками. При движении эндолимфы (во время угловых ускорений), когда волоски сгибаются в одну сторону, волосковые клетки возбуждаются, а при противоположно направленном движении — тормозятся. Это связано с тем, что механическое управление ионными каналами мембраны волоска с помощью микрофиламентов, описанное в разделе «механизмы слуховой рецепции», зависит от направления сгиба волоска: отклонение в одну сторону приводит к открыванию каналов и деполяризации волос- ковой клетки, а отклонение в противоположном направлении вызывает закрытие каналов и гиперполяризацию рецептора. В волос- ковых клетках преддверия и ампулы при их сгибании генерируется рецепторный потенциал, который усиливает выделение ацетилхо- лина и через синапсы активирует окончания волокон вестибулярного нерва.
Волокна вестибулярного нерва (отростки биполярных нейронов) направляются в продолговатый мозг. Импульсы, приходящие по этим волокнам, активируют нейроны бульбарного вестибулярного комплекса, в состав которого входят ядра: преддверное верхнее, или Бехтерева, преддверное латеральное, или Дейтерса, Швальбе и др. Отсюда сигналы направляются во многие отделы ЦНС: спинной мозг, мозжечок, глазодвигательные ядра, кору большого мозга, ретикулярную формацию и ганглии автономной нервной системы.
Электрические явления в вестибулярной системе. Даже в полном покое в вестибулярном нерве регистрируется спонтанная им- пульсация. Частота разрядов в нерве повышается при поворотах головы в одну сторону и тормозится при поворотах в другую (детекция направления движения). Реже частота разрядов повышается или, наоборот, тормозится при любом движении. У 2/з волокон обнаруживают эффект адаптации (уменьшение частоты разрядов) во время длящегося действия углового ускорения. Нейроны вестибулярных ядер обладают способностью реагировать и на изменение положения конечностей, повороты тела, сигналы от внутренних органов, т. е. осуществлять синтез информации, поступающей из разных источников.
Комплексные рефлексы, связанные с вестибулярной стимуляцией. Нейроны вестибулярных ядер обеспечивают контроль и управление различными двигательными реакциями. Важнейшими из этих реакций являются следующие: вестибулоспинальные, вести- буловегетативные и вестибулоглазодвигательные. Вестибулоспинальные влияния через вестибуло-, ретикуло- и руброспинальные тракты изменяют импульсацию нейронов сегментарных уровней спинного мозга. Так осуществляется динамическое перераспределение тонуса скелетной мускулатуры и включаются рефлекторные реакции, необходимые для сохранения равновесия. Мозжечок при этом ответствен за фазический характер этих реакций: после его удаления вестибулоспинальные влияния становятся по преимуществу тоническими. Во время произвольных движений вестибулярные влияния на спинной мозг ослабляются.
В вестибуловегетативные реакции вовлекаются сердечно-сосудистая система, пищеварительный тракт и другие внутренние органы. При сильных и длительных нагрузках на вестибулярный аппарат возникает патологический симптомокомплекс, названный болезнью движения, например морская болезнь. Она проявляется изменением сердечного ритма (учащение, а затем замедление), сужением, а затем расширением сосудов, усилением сокращений желудка, головокружением, тошнотой и рвотой. Повышенная склонность к болезни движения может быть уменьшена специальной тренировкой (вращение, качели) и применением ряда лекарственных средств.
Вестибулоглазодвигательные рефлексы (глазной нистагм) состоят в медленном движении глаз в противоположную вращению сторону, сменяющемся скачком глаз обратно. Само возникновение и характеристика вращательного глазного нистагма — важные показатели состояния вестибулярной системы, они широко используются в морской, авиационной и космической медицине, а также в эксперименте и клинике.
Основные афферентные пути и проекции вестибулярных сигналов. Есть два основных пути поступления вестибулярных сигналов в кору большого мозга: прямой — через дорсомедиальную часть вентрального постлатерального ядра и непрямой вестибулоцере- беллоталамический путь через медиальную часть вентролатераль- ного ядра. В коре полушарий большого мозга основные афферентные проекции вестибулярного аппарата локализованы в задней части постцентральной извилины. В моторной зоне коры спереди от нижней части центральной борозды обнаружена вторая вестибулярная зона.
Функции вестибулярной системы. Вестибулярная система помогает организму ориентироваться в пространстве при активном и пассивном движении. При пассивном движении корковые отделы системы запоминают направление движения, повороты и пройден-
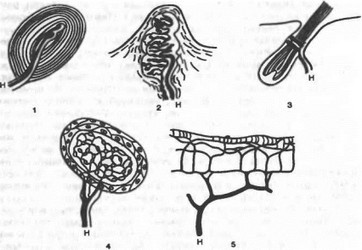
Рис. 14.19. Виды кожных рецепторов.
1—пластинчатое тельце (Фатера — Пачини); 2 — осязательное тельце (Мейсснера); 3 — нервное сплетение в волосяном мешочке; 4 — луковица (колба Краузе); 5 — нервное сплетение в роговой оболочке; Н — нервное волокно.
ное расстояние. Следует подчеркнуть, что в нормальных условиях пространственная ориентировка обеспечивается совместной деятельностью зрительной и вестибулярной систем. Чувствительность вестибулярной системы здорового человека очень высока: отолитовый аппарат позволяет воспринять ускорение прямолинейного движения, равное всего 2 см/с2. Порог различения наклона головы в сторону — всего около 1°, а вперед и назад — 1,5—2°. Рецепторная система полукружных каналов позволяет человеку замечать ускорения вращения 2—3° • с-2.