
У млекопитающих известно пять классов иммуноглобулинов: IgM, IgG, IgA, IgE и IgD, которые имеют общий план строения, но отличаются структурными особенностями тяжелых (Н) цепей.
Первый шаг к пониманию строения иммуноглобулинов был сделан английским исследователем Р.Портером в 1959 г. Он продемонстрировал, что обработка кроличьих антител IgG-класса ферментом папаином расщепляет молекулу на два основных фрагмента с мол. массами 45 кД и 50 кД. Один из этих фрагментом сохранял способность связывать антиген и в силу этого получил название Fab-фрагмента (от англ, "antigen binding”). Второй фрагмент не взаимодействовал с антигеном. Его удалось легко кристаллизовать, что и послужило основанием для его обозначения как Fc-фрагмента (от англ, “ciystallizable”). В количественном отно
шении Fab-фрагментов в два раза больше, чем Fc-фрагментов. Естественно было предположить, что молекула IgG имеет два участка, которые взаимодействуют с антигеном, и один участок — антигенноинертный. Выяснено, что папаин разрушает иммуноглобулин в шарнирной области, выше межцепьевых, дисульфидных связей, что и приводит к образованию двух идентичных и одного отличающегося участков (рис. 2.3.).
Опыты Ниссонова подтвердили это предположение. При работе с пепсином выделен один двухвалентный, антигенсвязываю- щий фрагмент. Часть молекулы IgG, соответствующая Fc-фрагменту, полностью разрушается. Получение двухвалентного фрагмента иммуноглобулина обеспечено действием пепсина на дистальный конец шарнирной области. В результате N-концевая половина молекулы остается нетронутой. Такой двухвалентный фрагмент обозначают как F(ab)2.
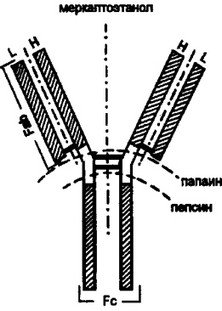
Рис. 2.3. Определение структуры антител с помощью цротеолитических фермеп-
тов.
Папаин разрушает молекулу IgG в шарнирной области (незапггрихованная часть рисунка) выше дисульфидных связей. В результате образуется два основных фрагмента с мол. массами 45 и 50 кД. Один из этих фрагментов (Fab) сохраняет способность связывать антиген. Другой С-концевой фрагмент (Fc) не обладает такой способностью. В молярном отношении Fab-фрагментов в два раза больше, чем Fc-фрагментов. При использовании пепсина расщепление молекулы иммуноглобулина происходит в дистальной части шарнирной области, что приводит к получению фрагментов с мол. массой более 90 кД. С-концевая часть молекулы разрушается. Полученный фрагмент способен связывать две молекулы антигена и в силу этого получил обозначение F(ab)2. С помощью меркаптоэтанола, разрушающего дисульфидные (-S-S-) связи, установлено наличие в структуре иммуноглобулинов двух тяжелых (Н) и двух легких (L) цепей. Линии “пунктир” и “пунктир- точка” указывают на места разрыва молекулы
И, наконец, исследования Эдельмана, выполненные с использованием меркаптоэтанола и ряда других соединений, которые разрушают межцепьевые -S-S- связи, показали наличие в молекуле иммуноглобулина двух тяжелых (Н) цепей с мол. массой каждой из них около 50 кД и двух легких (L) с мол. массой 25 кД.
Для получения информации о строении и молекулярных основах специфичности антител необходимо было иметь значительное количество полностью идентичных иммуноглобулинов. Исследования с сывороточными антителами от нормальных доноров не давали такой возможности, так как подобные антитела, являясь производными нескольких клеточных клонов, могли варьировать по тонкой специфичности антигенсвязывающего центра и, кроме того, относиться к различным классам иммуноглобулинов. Необходима была экспериментальная модель, позволяющая работать с иммуноглобулинами, продуцируемыми одним клоном клеток и в силу этого представляющими собой полностью идентичные молекулы. Такой моделью являются злокачественно трансформированные плазматические клетки больных миеломой. Больные данной формой рака имеют около 95% иммуноглобулинов сыворотки крови, относящихся к одному клону — потомству одной исходной клетки. К клональным белкам относятся и белки Бенс-Джонса, названные по имени первооткрывателя. Эти белки представляют собой димеры легких цепей иммуноглобулинов (L-L-цепи). Они обнаруживаются в большом количестве в моче больных миеломой и также являются удобным объектом для секвенирования. У мышей можно индуцировать миелому минеральными маслами. Существует большой набор клонированных плазмоцитом, продуцирующих соответствующие миеломные белки — иммуноглобулины. Эти линии клеток получили название MOPC’s (от англ, “mineral oil induction plasmacytome cells”).
Изучение полной аминокислотной последовательности различных миеломных белков выявило принципиальные особенности в строении иммуноглобулинов. Иммуноглобулины разных классов характеризуются общим планом строения. На рис. 2.4 представлена схема организации IgG. Этот иммуноглобулин содержит две тяжелые (Н) цепи и две легкие (L) цепи, которые объединены в четырехцепочечную молекулу посредством ковалентных, межцепьевых, дисульфидных связей (-S-S-). Каждая цепь включает вариабельную область (соответственно VL и VH для V- и Н- цепей), от которой зависит специфичность иммуноглобулинов как антител, и константную (С), подразделяющуюся на гомологичные ;тастки: Сн1, ^н2, Сн3. L-цепь имеет один константный участок (С^. Между СН1 и Сн2 расположена так называемая шарнирная область, обогащенная пролиновыми остатками. Повышенное
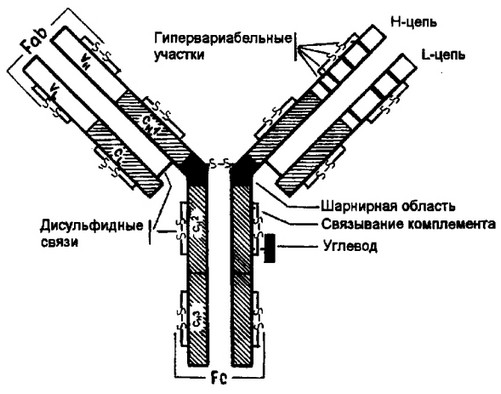
Рис. 2.4. Структура IgG.
Иммуноглобулин G содержит две тяжелые (Н) цепи с мол. массой 50 кД и две легкие (L) с мол. массой 25 кД, которые объединены в единую молекулу с помощью ковалентных дисульфидных связей -S-S-. Каждая цепь содержит вариабельную область (VL и VH для L- и Н-цепей соответственно) и константную (С) область, подразделяющуюся у Н-цепей на гомологичные участки (домены): CHlf Сн2, Сн3. L-цепь имеет один константный участок — CL. Dr взаимодействия VH- и Уь-областей зависит специфичность иммуношобулинов как антител. В аминокислотной последовательности V-доменов имеются положения, характеризующиеся частой заменой аминокислот от белка к белку (гилервариабельные участки) и более консервативные положения. Между Сн1- и Сн2-доменами Н-цепи находится шарнирная область, от которой зависит подвижность Fab-фрагмента. С^-до- мен является местом присоединения углеводов и связывания комплемента. Сн3- домен взаимодействует с Fc-рецептором на поверхности клеток, принимающих участие в иммунологических реакциях
содержание пролива в данной области обеспечивает конформа- ционную гибкость молекулы, что необходимо для лучшего взаимодействия с антигенными детерминантами, более выраженными на поверхности клеток.
Впервые в 1969 г., еще до получения рентгеноструктурных данных, Дж. Эдельман предположил, что каждый гомологичный участок организован в замкнутую сферу — домен, за счет внутри- цепьевых дисульфидных связей, образующихся полуцистеивовыми остатками. Дисульфидная связь замыкает в петлю около 60 аминокислот. Приблизительно по 20 аминокислот, не входящих в замкнутую часть участка, служат для взаимодействия с соседними доменами (рис. 2.5). Рентгенострукгурный анализ подтвердил общий 56
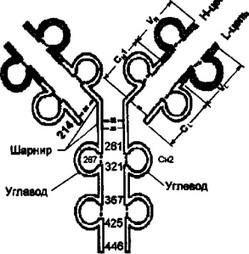
Рис. 2.S. Принцип доменной организации иммуноглобулинов.
Представлена схема доменной организации IgG. Каждый домен включает приблизительно 100-110 аминокислотных остатков. Около 60 остатков оказываются заключенными в петлю дисульфидной -S-S- связью. По 20 аминокислотных остатков, не входящих в замкнутую часть домена, служат для соединения с соседними доменами. Цифры обозначают последовательность аминокислотных остатков в полипептидах
принцип доменной организации полипептидных цепей иммуноглобулинов и вскрыл ряд тонких деталей строения. На рис. 2.6 представлено схематическое изображение пространственной организации V- и С-доменов легкой цепи. Домен состоит из двух слоев с Р-складчатой структурой, один из которых построен из четырех
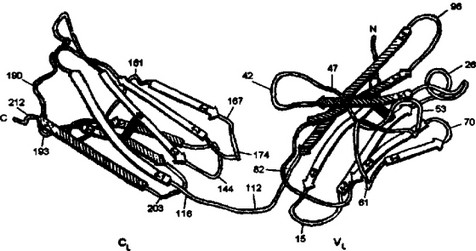
Рис. 2.6. Структурная организация легкой цепи (VL- и CL- доменов).
Организация доменов воспроизведена на основании данных по рентгеноструктурному анализу. Стрелки показывают участки с антипараллельной бета-струк- турной трехсегментного (заштрихован) и четырехсегментного (незаштрихован) слоев. Цифрами обозначены положения аминокислотных остатков. Черные линии — дисульфидаые соединения
антипараллельных сегментов, а второй — из трех сегментов. Слои ковалентно связаны дисульфидным мостиком примерно в середине домена. Общая пространственная организация IgG человека представлена на рис. 2.7. Видно, что тяжелые и легкие цепи, взаимодействуя друг с другом, образуют плотно упакованную структуру с тремя частями: два Fab-фрагмента и один Fc-фрагмент.
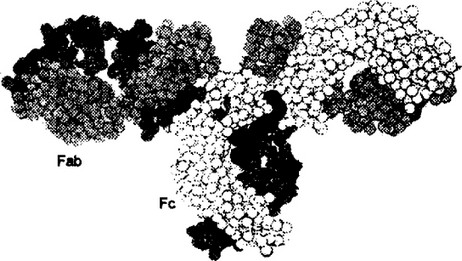
Рис. 2.7. Трехмерная структура IgG человека.
Светлое и темно-серое изображение обозначают тяжелые цепи; светло-серое — легкие цепи; черное — углеводы
Первый шаг к пониманию строения иммуноглобулинов был сделан английским исследователем Р.Портером в 1959 г. Он продемонстрировал, что обработка кроличьих антител IgG-класса ферментом папаином расщепляет молекулу на два основных фрагмента с мол. массами 45 кД и 50 кД. Один из этих фрагментом сохранял способность связывать антиген и в силу этого получил название Fab-фрагмента (от англ, "antigen binding”). Второй фрагмент не взаимодействовал с антигеном. Его удалось легко кристаллизовать, что и послужило основанием для его обозначения как Fc-фрагмента (от англ, “ciystallizable”). В количественном отно
шении Fab-фрагментов в два раза больше, чем Fc-фрагментов. Естественно было предположить, что молекула IgG имеет два участка, которые взаимодействуют с антигеном, и один участок — антигенноинертный. Выяснено, что папаин разрушает иммуноглобулин в шарнирной области, выше межцепьевых, дисульфидных связей, что и приводит к образованию двух идентичных и одного отличающегося участков (рис. 2.3.).
Опыты Ниссонова подтвердили это предположение. При работе с пепсином выделен один двухвалентный, антигенсвязываю- щий фрагмент. Часть молекулы IgG, соответствующая Fc-фрагменту, полностью разрушается. Получение двухвалентного фрагмента иммуноглобулина обеспечено действием пепсина на дистальный конец шарнирной области. В результате N-концевая половина молекулы остается нетронутой. Такой двухвалентный фрагмент обозначают как F(ab)2.
Рис. 2.3. Определение структуры антител с помощью цротеолитических фермеп-
тов.
Папаин разрушает молекулу IgG в шарнирной области (незапггрихованная часть рисунка) выше дисульфидных связей. В результате образуется два основных фрагмента с мол. массами 45 и 50 кД. Один из этих фрагментов (Fab) сохраняет способность связывать антиген. Другой С-концевой фрагмент (Fc) не обладает такой способностью. В молярном отношении Fab-фрагментов в два раза больше, чем Fc-фрагментов. При использовании пепсина расщепление молекулы иммуноглобулина происходит в дистальной части шарнирной области, что приводит к получению фрагментов с мол. массой более 90 кД. С-концевая часть молекулы разрушается. Полученный фрагмент способен связывать две молекулы антигена и в силу этого получил обозначение F(ab)2. С помощью меркаптоэтанола, разрушающего дисульфидные (-S-S-) связи, установлено наличие в структуре иммуноглобулинов двух тяжелых (Н) и двух легких (L) цепей. Линии “пунктир” и “пунктир- точка” указывают на места разрыва молекулы
И, наконец, исследования Эдельмана, выполненные с использованием меркаптоэтанола и ряда других соединений, которые разрушают межцепьевые -S-S- связи, показали наличие в молекуле иммуноглобулина двух тяжелых (Н) цепей с мол. массой каждой из них около 50 кД и двух легких (L) с мол. массой 25 кД.
Для получения информации о строении и молекулярных основах специфичности антител необходимо было иметь значительное количество полностью идентичных иммуноглобулинов. Исследования с сывороточными антителами от нормальных доноров не давали такой возможности, так как подобные антитела, являясь производными нескольких клеточных клонов, могли варьировать по тонкой специфичности антигенсвязывающего центра и, кроме того, относиться к различным классам иммуноглобулинов. Необходима была экспериментальная модель, позволяющая работать с иммуноглобулинами, продуцируемыми одним клоном клеток и в силу этого представляющими собой полностью идентичные молекулы. Такой моделью являются злокачественно трансформированные плазматические клетки больных миеломой. Больные данной формой рака имеют около 95% иммуноглобулинов сыворотки крови, относящихся к одному клону — потомству одной исходной клетки. К клональным белкам относятся и белки Бенс-Джонса, названные по имени первооткрывателя. Эти белки представляют собой димеры легких цепей иммуноглобулинов (L-L-цепи). Они обнаруживаются в большом количестве в моче больных миеломой и также являются удобным объектом для секвенирования. У мышей можно индуцировать миелому минеральными маслами. Существует большой набор клонированных плазмоцитом, продуцирующих соответствующие миеломные белки — иммуноглобулины. Эти линии клеток получили название MOPC’s (от англ, “mineral oil induction plasmacytome cells”).
Изучение полной аминокислотной последовательности различных миеломных белков выявило принципиальные особенности в строении иммуноглобулинов. Иммуноглобулины разных классов характеризуются общим планом строения. На рис. 2.4 представлена схема организации IgG. Этот иммуноглобулин содержит две тяжелые (Н) цепи и две легкие (L) цепи, которые объединены в четырехцепочечную молекулу посредством ковалентных, межцепьевых, дисульфидных связей (-S-S-). Каждая цепь включает вариабельную область (соответственно VL и VH для V- и Н- цепей), от которой зависит специфичность иммуноглобулинов как антител, и константную (С), подразделяющуюся на гомологичные ;тастки: Сн1, ^н2, Сн3. L-цепь имеет один константный участок (С^. Между СН1 и Сн2 расположена так называемая шарнирная область, обогащенная пролиновыми остатками. Повышенное
Рис. 2.4. Структура IgG.
Иммуноглобулин G содержит две тяжелые (Н) цепи с мол. массой 50 кД и две легкие (L) с мол. массой 25 кД, которые объединены в единую молекулу с помощью ковалентных дисульфидных связей -S-S-. Каждая цепь содержит вариабельную область (VL и VH для L- и Н-цепей соответственно) и константную (С) область, подразделяющуюся у Н-цепей на гомологичные участки (домены): CHlf Сн2, Сн3. L-цепь имеет один константный участок — CL. Dr взаимодействия VH- и Уь-областей зависит специфичность иммуношобулинов как антител. В аминокислотной последовательности V-доменов имеются положения, характеризующиеся частой заменой аминокислот от белка к белку (гилервариабельные участки) и более консервативные положения. Между Сн1- и Сн2-доменами Н-цепи находится шарнирная область, от которой зависит подвижность Fab-фрагмента. С^-до- мен является местом присоединения углеводов и связывания комплемента. Сн3- домен взаимодействует с Fc-рецептором на поверхности клеток, принимающих участие в иммунологических реакциях
содержание пролива в данной области обеспечивает конформа- ционную гибкость молекулы, что необходимо для лучшего взаимодействия с антигенными детерминантами, более выраженными на поверхности клеток.
Впервые в 1969 г., еще до получения рентгеноструктурных данных, Дж. Эдельман предположил, что каждый гомологичный участок организован в замкнутую сферу — домен, за счет внутри- цепьевых дисульфидных связей, образующихся полуцистеивовыми остатками. Дисульфидная связь замыкает в петлю около 60 аминокислот. Приблизительно по 20 аминокислот, не входящих в замкнутую часть участка, служат для взаимодействия с соседними доменами (рис. 2.5). Рентгенострукгурный анализ подтвердил общий 56
Рис. 2.S. Принцип доменной организации иммуноглобулинов.
Представлена схема доменной организации IgG. Каждый домен включает приблизительно 100-110 аминокислотных остатков. Около 60 остатков оказываются заключенными в петлю дисульфидной -S-S- связью. По 20 аминокислотных остатков, не входящих в замкнутую часть домена, служат для соединения с соседними доменами. Цифры обозначают последовательность аминокислотных остатков в полипептидах
принцип доменной организации полипептидных цепей иммуноглобулинов и вскрыл ряд тонких деталей строения. На рис. 2.6 представлено схематическое изображение пространственной организации V- и С-доменов легкой цепи. Домен состоит из двух слоев с Р-складчатой структурой, один из которых построен из четырех
Рис. 2.6. Структурная организация легкой цепи (VL- и CL- доменов).
Организация доменов воспроизведена на основании данных по рентгеноструктурному анализу. Стрелки показывают участки с антипараллельной бета-струк- турной трехсегментного (заштрихован) и четырехсегментного (незаштрихован) слоев. Цифрами обозначены положения аминокислотных остатков. Черные линии — дисульфидаые соединения
антипараллельных сегментов, а второй — из трех сегментов. Слои ковалентно связаны дисульфидным мостиком примерно в середине домена. Общая пространственная организация IgG человека представлена на рис. 2.7. Видно, что тяжелые и легкие цепи, взаимодействуя друг с другом, образуют плотно упакованную структуру с тремя частями: два Fab-фрагмента и один Fc-фрагмент.
Рис. 2.7. Трехмерная структура IgG человека.
Светлое и темно-серое изображение обозначают тяжелые цепи; светло-серое — легкие цепи; черное — углеводы