
Проприоцепторы, как уже говорилось, расположены в скелетных мышцах, суставных сумках и внутреннем ухе.
Мышечные веретена и тельца Гольджи. Проприоцепторы, находящиеся в мышцах, заключены в веретенах — группах особых мышечных волокон длиной 3—4 мм, окруженных капсу-
Сокращение мышцы
Растяжение мы*шцы
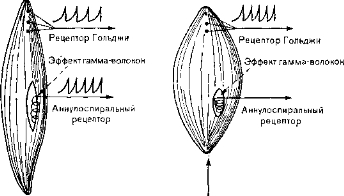
Рис. 24. Схема активации проприоцепторов. Аннулоспнральный рецептор разряжается только при растяжении мышцы, а рецептор Гольджи, с более высоким порогом, отвечает и на растяжение, и на сокращение. Механизм гамма-эфферентов контролирует только аннулоспиральные рецепторы.
лой. Вокруг таких волокон обвиваются аннулоспиральные рецепторы. При растяжении мышцы импульсы от рецепторов направляются к нейронам в сенсорном ганглии, расположенном около спинного мозга. В мышечном веретене есть и другие, менее важные рецепторы. В сухожилиях в месте их соединения с мышцами находятся рецепторы еще одного типа — тельца
Гольджи. Импульсы в них возникают, когда мышца растянута или сокращена. Волокна от телец Гольджи тоже идут к сенсорному ганглию, лежащему около спинного мозга. Порог раздражения у мышечных веретен гораздо ниже, чем у сухожильных телец Гольджи. Для возбуждения этих последних требуется
 чрезвычайно сильное растяжение или сокращение (рис. 24). В чувствительных аннулоспиральных рецепторах импульсы возникают только в ответ на растяжение мышцы, в которой они заключены. Импульсы, как правило, направляются самым коротким сенсорным путем к спинному мозгу и возбуждают так называемый рефлекс на растяжение. Функция этой системы состоит в том, чтобы мышечные волокна принимали и сохраняли такую длину, какая требуется для данного положения тела. Стимулом для этого рефлекса служит земное притяжение, которое нужно преодолевать для поддержания нужной позы.. Эти рефлексы называют также антигравитационными спинальными рефлексами. Импульсы не доходят до головного мозга, и рефлексы сохраняются у экспериментальных животных, лишенных всех нервных структур выше спинного мозга. Второй путь, по которому передаются проприо- цептивные импульсы, несущие важную статическую и кинетическую информацию, идет к большому мозгу и мозжечку.
чрезвычайно сильное растяжение или сокращение (рис. 24). В чувствительных аннулоспиральных рецепторах импульсы возникают только в ответ на растяжение мышцы, в которой они заключены. Импульсы, как правило, направляются самым коротким сенсорным путем к спинному мозгу и возбуждают так называемый рефлекс на растяжение. Функция этой системы состоит в том, чтобы мышечные волокна принимали и сохраняли такую длину, какая требуется для данного положения тела. Стимулом для этого рефлекса служит земное притяжение, которое нужно преодолевать для поддержания нужной позы.. Эти рефлексы называют также антигравитационными спинальными рефлексами. Импульсы не доходят до головного мозга, и рефлексы сохраняются у экспериментальных животных, лишенных всех нервных структур выше спинного мозга. Второй путь, по которому передаются проприо- цептивные импульсы, несущие важную статическую и кинетическую информацию, идет к большому мозгу и мозжечку.
Лабиринт. Лабиринт, или вестибулярный аппарат внутреннего уха, является вторым важным проприоцептором. Он структурно и генетически не зависит от улитки, хотя лежит во внутреннем ухе непосредственно над слуховым аппаратом. Вестибулярный аппарат состоит из отолитовых органов (круглого и
овального мешочков) и трех полукружных каналов, которые расположены в трех взаимно перпендикулярных плоскостях. Как и улитка, полукружные каналы заполнены эндолимфой. Рецепторными клетками, так же как и в кортиевом органе, являются волосковые клетки, погруженные в студенистую массу (рис. 25). Группы волосковых клеток найдены как в ампулах полукружных каналов, так и в отолитовых органах. Таким образом, каждый вестибулярный аппарат включает пять рецепторных систем. Волосковые клетки круглого и овального мешочков служат для получения информации о статическом положении головы. В студенистую массу погружены известковые частицы (отолиты), на которые воздействует сила тяжести. Порог раздражения этих рецепторов очень низок, поэтому импульсы от них постоянно идут в продолговатый мозг. Нервные окончания у основания волосковых клеток реагируют, когда известковые частички тянут волоски. Рецепторы полукружных каналов (волосковые клетки на кристах) воспринимают ускорение— положительное или отрицательное — движения головы. Таким образом, пока голова неподвижна, о ее положении сигнализируют в центр отолитовые органы, а о движениях головы сообщают рецепторы полукружных каналов. Порог раздражения полукружных каналов, реагирующих на изменения тока жидкости в их полости, выше, чем у отолитовых органов. Если голова неподвижна или движется с равномерной скоростью, эти рецепторы не посылают сигналов.
Не вся информация, передаваемая вестибулярным аппаратом, достигает коры. Часть ее доходит только до продолговатого или среднего мозга, приводя в действие сложный, но бессознательный автоматизм, регулирующий положение тела.
Корковое представительство. Проекционные зоны проприоце- пторов в коре изучались методом вызванных потенциалов. Как выяснилось, проприоцепторы имеют свое представительство главным образом в мозжечке, который служит важным центром координации движений, но группы клеток, отвечающих на стимуляцию различных проприоцепторов (в мышцах или в вестибулярном аппарате), найдены также в соматосенсорной области коры (поля 1, 2 и 3 по Бродману) (рис. 26). Поскольку нейроны в соматосенсорной коре расположены в соответствии с топографией тела (представительство головы и верхней части туловища находится в нижнем конце, близ сильвиевой борозды, а представительство ног — наверху), различные группы мышц тоже представлены в соответствующих частях соматосенсорной коры. Вестибулярный аппарат проецируется на представительство головы. Подобно зрительным и слуховым импульсам, сигналы, посылаемые проприоцепторамп, тоже имеют станцию переключения в таламусе. Таламическое ядро
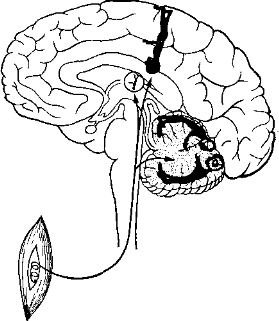
Рис. 26. Двойное представительство проприоцепции в коре большого мозга и в мозжечке (сильно упрощенная схема).
п. ventralis posterior служит промежуточной станцией для всех кожных, мышечных и висцеральных сенсорных путей.
Таким образом, проприоцептивные импульсы идут по сложному, самовозбуждающемуся кольцевому пути, который включает сенсорные и моторные зоны коры, определенные группы нейронов в стволе мозга (анатомы назвали их базальными ганглиями), упомянутые выше нейроны таламуса и мозжечковые структуры. Эту реверберационную систему, которая регулирует позу и координирует движения, неврологи называют экстрапирамидной системой. В то время как пирамидный тракт, идущий от моторной коры к мотонейронам спинного мозга, состоит всего из двух или трех нейронов и осуществляет точные, запланированные произвольные движения, экстрапира- мидный тракт управляет бессознательными, грубыми движениями. Рецепторами, снабжающими эту систему информацией, служат проприоцепторы, о которых шла речь выше.