
Принимая во внимание важность процессов гаметогенеза и начальных этапов эмбриогенеза в этиологии и патогенезе хромосомной патологии, в данном разделе будут рассмотрены: гаметогенез (сперматогенез и оогенез) и оплодотворение.
Все половые клетки млекопитающих и человека берут начало от первичных половых клеток (ПИК) — гоноцитов. Происхождение ППК до настоящего времени окончательно не выяснено. Не вызывает, однако, сомнения, что эти клетки возникают значительно раньше, чем появляются зачатки гонад, то есть они имеют экстрагонадное происхождение. Согласно существующим представлениям ППК могут быть обнаружены в первичной полоске уже на 16—18-й день развития, затем они перемещаются в желточную (внезародышевую) энтодерму у основания аллантоиса, мигрируют в энтодерму средней кишки, откуда и попадают в половые валики — зачатки гонад [54, 56, 172]. В последнее время получены данные о том, что ППК выделяются в самостоятельный эмбриональный зачаток значительно раньше, еще во время дробления и формирования бластоцисты.
Попав в зачатки гонад, гоноциты впервые обнаруживают признаки полового диморфизма. При формировании мужских гонад (семенников) они окружаются клетками целомического эпителия, образуя так называемые «половые тяжи», в составе которых пребывают в латентном, недифференцированном состоянии (сперматогонии) вплоть до начала полового созревания.
При формировании женских гонад (яичников) гоноциты задерживаются в наружном, корковом слое мезенхимной ткани половых валиков, активно пролиферируют, вступают в мейоз, после чего каждый из них окружается фолликулярными клетками и, в виде ооцитов 1-го порядка, сохраняется до полового созревания. Принципиальная схема гаметогенеза у млекопитающих и человека приведена на рисунке 1.1.
Общая продолжительность сперматогенеза у человека составляет 72 дня. За это время стволовые клетки сперматогенного ряда (спер- матогонии), находящиеся в глубине извитых семенных канальцев, проходят длительный путь дифференцировки до зрелых, практически лишенных цитоплазмы, сперматозоидов, содержащих гаплоидный набор хромосом. В процессе сперматогенеза различают две фазы — тестикулярную и эпидидемальную. Во время первой происходят основные этапы дифференцировки сперматогоний в сперматозоиды; во время второй завершается созревание спермиев. В результате накопления мукополисахаридов, холестерина, других защитных белков, меняются свойства наружных мембран, спермии приобретают подвижность.
Сперматогенез (тестикулярная фаза) включает два последовательных этапа: собственно сперматогенез и спермиогенез. Тестикулярная фаза контролируется гормонами гипофиза (фолликулостимулирующим
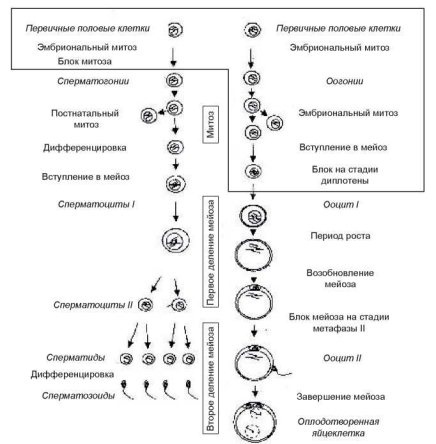
Рис. 1.1. Принципиальная схема сперматогенеза (слева) и оогенеза (справа) у млекопитающих и человека [895]
и лютеотропным) и собственными гормонами семенников — тестикулярными андрогенами (тестостероном, андростендионом и другими), которые продуцируются клетками Лейдига, находящимися в строме извитых семенных канальцев.
На 1-м этапе вступающие в мейоз клетки (сперматоциты 1-го порядка) претерпевают два последовательных мейотических деления. При этом из одного сперматоцита 1-го порядка возникают 4 клетки (сперматиды) с гаплоидным числом хромосом (рис. 1.1). Все процессы дифференцировки проходят в стенке извитых семенных канальцев. При этом клетки сперматогенного ряда находятся непосредственно в цитоплазме клеток Сертоли, которые обеспечивают питание сперма- тоцитов и сперматид.
Во время спермиогенеза гаплоидные клетки — сперматиды — проходят ряд последовательных стадий дифференцировки (фаза Гольджи, фаза колпачка, акросомная фаза, фаза созревания). Они утрачивают цитоплазму, формируют специальные органоиды (хвост, шейку, ак- росому) (рис. 1.2). Акросома возникает непосредственно из мембран аппарата Гольджи, покрывает в виде колпачка переднюю часть головки спермия (примерно до ее середины) и содержит набор литических лизосомных ферментов, важных для оплодотворения (рис. 1.2).
Особенно существенные изменения происходят непосредственно в ядре клеток. ДНК в составе хромосом утрачивает типичную для соматических клеток нуклеосомную организацию. Гистоновые белки, характерные для функционально активной ДНК, заменяются на кис-
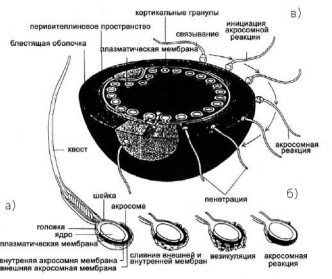
Рис. 1.2. Строение зрелого сперматозоида (а), акросомная реакция (б), последовательные этапы оплодотворения (в)
лые белки, богатые аргинином и протаминами. Спирализация ДНК достигает максимальной величины. Ежедневно у человека активного репродуктивного возраста продуцируется свыше 10 млн зрелых сперматозоидов.
В отличие от мужских половых клеток родоначальники женских половых клеток — оогонии — претерпевают важнейшие стадии диф- ференцировки, включая все этапы профазы мейоза (рис. 1.1), еще во внутриутробном периоде развития. Более подробно временные и цитогенетические характеристики мейоза у зародышей женского пола и у половозрелых женщин представлены на рисунке 1.3. Временные особенности основных периодов оогенеза у человека приведены в таблице 1.2. Достигнув зачатков будущих яичников (половых валиков) примерно к концу 1-го — середине 2-го месяца беременности, гоноци- ты теряют амебоидную подвижность, вступают в контакт с клетками фолликулярного эпителия и преобразуются в оогонии. В течение пос-
Таблица 1.2. Временные особенности основных периодов оогенеза у разных млекопитающих и человека [52]
Примечание. Периоды оогенеза даны в соответствующих стадиях онтогенеза: д. б. — дни беременности, д. п. р. — дни после рождения, м. б. — месяцы беременности, м. п. р. — месяцы после рождения
ледующих 3-4 месяцев оогонии активно делятся митозом. В результате их число возрастает от исходных 1500-2000 клеток до нескольких миллионов. Максимальное число оогоний (до 7 млн) находится в яичниках плодов женского пола на 7-м месяце беременности. Сразу же за периодом размножения следует апоптоз — запрограммированная гибель большей части оогониев. Причины апоптоза остаются невыясненными. Неясна и селективная роль такой массовой клеточной гибели. Возможно, погибают те оогонии, которые по тем или иным причинам не могут трансформироваться в ооциты и вступить в мейоз, либо, что кажется более правдоподобным, гибнущие клетки — это ооциты, находящиеся в профазе мейоза.
Количество женских половых клеток к концу беременности и у новорожденных уменьшается в среднем до 2 млн, к 7 годам — до 300 000, а к началу полового созревания — до 40 000. Реально в течение всей жизни овулирует не более 400-500 ооцитов. Значительная часть естественной убыли женских половых клеток происходит в результате апоптоза оогониев, другие погибают уже внутри атретических фолликулов, которые не доходят до овуляции. Уместно также отметить, что в отличие от млекопитающих, процессы оогенеза у человека протекают асинхронно, а потому значительно растянуты во времени.
Уже с 3-го месяца беременности часть оогониев завершает циклы митотических делений, трансформируется в ооциты и вступает в период роста. Они увеличиваются в размерах, окружаются фолликулярными клетками, вступают в профазу мейоза (рис. 1.3). Однако в отличие от мужского мейоза, в оогенезе вслед за профазой не наступает метафаза, а мейоз блокируется, и ооциты надолго, вплоть до начала полового созревания, переходят в состояние покоя — диктиотену. Предполагается, что блокада мейоза связана с действием особых факторов, секретиру- емых соматическими (фолликулярными) клетками гонады [54]. Окруженные одним слоем фолликулярных клеток ооциты образуют так называемые первичные (примордиальные) фолликулы. До полового созревания длится период медленного роста, во время которого прогрессивно увеличивается число слоев фолликулярных клеток, окружающих ооцит на стадии покоя (диктиотены). Ядро ооцита на этой стадии очень крупное, светлое, называется иногда «зародышевым пузырьком». Характерной структурой такого ядра у человека являются
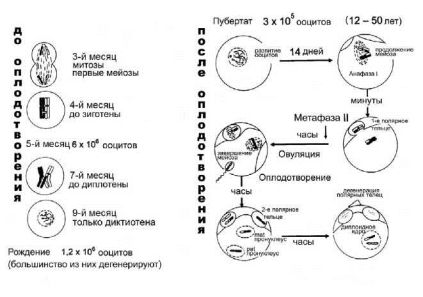
Рис. 1.3. Цитогенетические и временные характеристики профазы мейоза у плодов женского пола и стадий созревания яйцеклеток после полового созревания [804]
«ламповые щетки» — петли ДНК, на которых происходит активный синтез РНК-комплексов, откладывающихся в ооплазме до момента оплодотворения. Размеры ооцита по мере увеличения числа фолликулярных клеток также увеличиваются. Рост самого ооцита прекращается только с началом периода быстрого роста его фолликула, что совпадает с периодом полового созревания. В это время внутри фолликула образуется полость (антрум), которая заполняется жидкостью. Ее размеры быстро увеличиваются. Фолликул превращается в Граафов пузырек [52].
Созревание ооцитов начинается с возобновления мейоза и заканчивается только после оплодотворения, когда завершается 2-е мейо- тическое деление (рис. 1.3). С наступлением активного репродуктивного возраста ооциты группами (5-10 шт.) вступают в мейоз, однако в большинстве случаев в каждом цикле овулирует только один, наиболее продвинутый в развитии доминантный фолликул, тогда как ооциты
в остальных фолликулах, вступившие в период созревания, прекращают развитие и подвергаются атрезии.
Рост и созревание фолликулов с находящимися в них ооцитами находится под гормональным контролем как со стороны гипофиза (фолликулостимулирующий гормон — ФСГ, лютетропный гормон — ЛГ) и гипоталамуса (пролактин — гонадотропин-релизинг гормон), так и самого яичника (эстрогены, гормоны фолликулярных клеток, прогестерон — гормон желтого тела). При этом период роста ооцитов, особенно период быстрого роста, контролируется преимущественно ФСГ, а период созревания — ЛГ. Примерно за сутки до овуляции, то есть до разрыва Граафова пузырька и выхода ооцита, отмечается пик подъема ЛГ.
Контролирующие механизмы оогенеза и особенности гормональной регуляции этого процесса подробно рассмотрены в ряде обстоятельных монографий и обзоров [52, 54, 94, 129, 556].
Все половые клетки млекопитающих и человека берут начало от первичных половых клеток (ПИК) — гоноцитов. Происхождение ППК до настоящего времени окончательно не выяснено. Не вызывает, однако, сомнения, что эти клетки возникают значительно раньше, чем появляются зачатки гонад, то есть они имеют экстрагонадное происхождение. Согласно существующим представлениям ППК могут быть обнаружены в первичной полоске уже на 16—18-й день развития, затем они перемещаются в желточную (внезародышевую) энтодерму у основания аллантоиса, мигрируют в энтодерму средней кишки, откуда и попадают в половые валики — зачатки гонад [54, 56, 172]. В последнее время получены данные о том, что ППК выделяются в самостоятельный эмбриональный зачаток значительно раньше, еще во время дробления и формирования бластоцисты.
Попав в зачатки гонад, гоноциты впервые обнаруживают признаки полового диморфизма. При формировании мужских гонад (семенников) они окружаются клетками целомического эпителия, образуя так называемые «половые тяжи», в составе которых пребывают в латентном, недифференцированном состоянии (сперматогонии) вплоть до начала полового созревания.
При формировании женских гонад (яичников) гоноциты задерживаются в наружном, корковом слое мезенхимной ткани половых валиков, активно пролиферируют, вступают в мейоз, после чего каждый из них окружается фолликулярными клетками и, в виде ооцитов 1-го порядка, сохраняется до полового созревания. Принципиальная схема гаметогенеза у млекопитающих и человека приведена на рисунке 1.1.
- сперматогенез
Общая продолжительность сперматогенеза у человека составляет 72 дня. За это время стволовые клетки сперматогенного ряда (спер- матогонии), находящиеся в глубине извитых семенных канальцев, проходят длительный путь дифференцировки до зрелых, практически лишенных цитоплазмы, сперматозоидов, содержащих гаплоидный набор хромосом. В процессе сперматогенеза различают две фазы — тестикулярную и эпидидемальную. Во время первой происходят основные этапы дифференцировки сперматогоний в сперматозоиды; во время второй завершается созревание спермиев. В результате накопления мукополисахаридов, холестерина, других защитных белков, меняются свойства наружных мембран, спермии приобретают подвижность.
Сперматогенез (тестикулярная фаза) включает два последовательных этапа: собственно сперматогенез и спермиогенез. Тестикулярная фаза контролируется гормонами гипофиза (фолликулостимулирующим
Рис. 1.1. Принципиальная схема сперматогенеза (слева) и оогенеза (справа) у млекопитающих и человека [895]
и лютеотропным) и собственными гормонами семенников — тестикулярными андрогенами (тестостероном, андростендионом и другими), которые продуцируются клетками Лейдига, находящимися в строме извитых семенных канальцев.
На 1-м этапе вступающие в мейоз клетки (сперматоциты 1-го порядка) претерпевают два последовательных мейотических деления. При этом из одного сперматоцита 1-го порядка возникают 4 клетки (сперматиды) с гаплоидным числом хромосом (рис. 1.1). Все процессы дифференцировки проходят в стенке извитых семенных канальцев. При этом клетки сперматогенного ряда находятся непосредственно в цитоплазме клеток Сертоли, которые обеспечивают питание сперма- тоцитов и сперматид.
Во время спермиогенеза гаплоидные клетки — сперматиды — проходят ряд последовательных стадий дифференцировки (фаза Гольджи, фаза колпачка, акросомная фаза, фаза созревания). Они утрачивают цитоплазму, формируют специальные органоиды (хвост, шейку, ак- росому) (рис. 1.2). Акросома возникает непосредственно из мембран аппарата Гольджи, покрывает в виде колпачка переднюю часть головки спермия (примерно до ее середины) и содержит набор литических лизосомных ферментов, важных для оплодотворения (рис. 1.2).
Особенно существенные изменения происходят непосредственно в ядре клеток. ДНК в составе хромосом утрачивает типичную для соматических клеток нуклеосомную организацию. Гистоновые белки, характерные для функционально активной ДНК, заменяются на кис-
Рис. 1.2. Строение зрелого сперматозоида (а), акросомная реакция (б), последовательные этапы оплодотворения (в)
лые белки, богатые аргинином и протаминами. Спирализация ДНК достигает максимальной величины. Ежедневно у человека активного репродуктивного возраста продуцируется свыше 10 млн зрелых сперматозоидов.
- оогенез
В отличие от мужских половых клеток родоначальники женских половых клеток — оогонии — претерпевают важнейшие стадии диф- ференцировки, включая все этапы профазы мейоза (рис. 1.1), еще во внутриутробном периоде развития. Более подробно временные и цитогенетические характеристики мейоза у зародышей женского пола и у половозрелых женщин представлены на рисунке 1.3. Временные особенности основных периодов оогенеза у человека приведены в таблице 1.2. Достигнув зачатков будущих яичников (половых валиков) примерно к концу 1-го — середине 2-го месяца беременности, гоноци- ты теряют амебоидную подвижность, вступают в контакт с клетками фолликулярного эпителия и преобразуются в оогонии. В течение пос-
Таблица 1.2. Временные особенности основных периодов оогенеза у разных млекопитающих и человека [52]
|
Объект |
Вступление ооцитов в мейоз |
Размножение оогониев |
Рост ооцитов |
Созревание ооцитов, часы |
|
|
Профаза мейоза |
Диктиотена |
||||
|
Человек |
Асинхронное |
2-5 м. б. |
2,5-8 м. б. |
8-9 м. б. — 13,514 лет |
54-60 |
|
Макака |
» |
2-5 м. б. |
2-6 м. б. |
5 м. б. — 2-3 года |
54-60 |
|
Мышь |
Синхронное |
10-14 д. б. |
14-20 д. б. |
2-3 д. п. р. — 1,5 м. п. р. |
16-20 |
|
Крыса |
» |
14-17 д. б. |
17,5 д. б. — 4 д. п. р. |
5 д. п. р. — 1,5 м. п. р. |
16-20 |
|
Кролик |
» |
14-20 д. п. р. |
3-20 д. п. р. |
20 д. п. р. — 4-5 м. п. р. |
15-18 |
|
Золотистый хомячок |
» |
10-17 д. б. |
1-9 д. п. р. |
10 д. п. р. — 1,5 м. п. р. |
16-20 |
Примечание. Периоды оогенеза даны в соответствующих стадиях онтогенеза: д. б. — дни беременности, д. п. р. — дни после рождения, м. б. — месяцы беременности, м. п. р. — месяцы после рождения
ледующих 3-4 месяцев оогонии активно делятся митозом. В результате их число возрастает от исходных 1500-2000 клеток до нескольких миллионов. Максимальное число оогоний (до 7 млн) находится в яичниках плодов женского пола на 7-м месяце беременности. Сразу же за периодом размножения следует апоптоз — запрограммированная гибель большей части оогониев. Причины апоптоза остаются невыясненными. Неясна и селективная роль такой массовой клеточной гибели. Возможно, погибают те оогонии, которые по тем или иным причинам не могут трансформироваться в ооциты и вступить в мейоз, либо, что кажется более правдоподобным, гибнущие клетки — это ооциты, находящиеся в профазе мейоза.
Количество женских половых клеток к концу беременности и у новорожденных уменьшается в среднем до 2 млн, к 7 годам — до 300 000, а к началу полового созревания — до 40 000. Реально в течение всей жизни овулирует не более 400-500 ооцитов. Значительная часть естественной убыли женских половых клеток происходит в результате апоптоза оогониев, другие погибают уже внутри атретических фолликулов, которые не доходят до овуляции. Уместно также отметить, что в отличие от млекопитающих, процессы оогенеза у человека протекают асинхронно, а потому значительно растянуты во времени.
Уже с 3-го месяца беременности часть оогониев завершает циклы митотических делений, трансформируется в ооциты и вступает в период роста. Они увеличиваются в размерах, окружаются фолликулярными клетками, вступают в профазу мейоза (рис. 1.3). Однако в отличие от мужского мейоза, в оогенезе вслед за профазой не наступает метафаза, а мейоз блокируется, и ооциты надолго, вплоть до начала полового созревания, переходят в состояние покоя — диктиотену. Предполагается, что блокада мейоза связана с действием особых факторов, секретиру- емых соматическими (фолликулярными) клетками гонады [54]. Окруженные одним слоем фолликулярных клеток ооциты образуют так называемые первичные (примордиальные) фолликулы. До полового созревания длится период медленного роста, во время которого прогрессивно увеличивается число слоев фолликулярных клеток, окружающих ооцит на стадии покоя (диктиотены). Ядро ооцита на этой стадии очень крупное, светлое, называется иногда «зародышевым пузырьком». Характерной структурой такого ядра у человека являются
Рис. 1.3. Цитогенетические и временные характеристики профазы мейоза у плодов женского пола и стадий созревания яйцеклеток после полового созревания [804]
«ламповые щетки» — петли ДНК, на которых происходит активный синтез РНК-комплексов, откладывающихся в ооплазме до момента оплодотворения. Размеры ооцита по мере увеличения числа фолликулярных клеток также увеличиваются. Рост самого ооцита прекращается только с началом периода быстрого роста его фолликула, что совпадает с периодом полового созревания. В это время внутри фолликула образуется полость (антрум), которая заполняется жидкостью. Ее размеры быстро увеличиваются. Фолликул превращается в Граафов пузырек [52].
Созревание ооцитов начинается с возобновления мейоза и заканчивается только после оплодотворения, когда завершается 2-е мейо- тическое деление (рис. 1.3). С наступлением активного репродуктивного возраста ооциты группами (5-10 шт.) вступают в мейоз, однако в большинстве случаев в каждом цикле овулирует только один, наиболее продвинутый в развитии доминантный фолликул, тогда как ооциты
в остальных фолликулах, вступившие в период созревания, прекращают развитие и подвергаются атрезии.
Рост и созревание фолликулов с находящимися в них ооцитами находится под гормональным контролем как со стороны гипофиза (фолликулостимулирующий гормон — ФСГ, лютетропный гормон — ЛГ) и гипоталамуса (пролактин — гонадотропин-релизинг гормон), так и самого яичника (эстрогены, гормоны фолликулярных клеток, прогестерон — гормон желтого тела). При этом период роста ооцитов, особенно период быстрого роста, контролируется преимущественно ФСГ, а период созревания — ЛГ. Примерно за сутки до овуляции, то есть до разрыва Граафова пузырька и выхода ооцита, отмечается пик подъема ЛГ.
Контролирующие механизмы оогенеза и особенности гормональной регуляции этого процесса подробно рассмотрены в ряде обстоятельных монографий и обзоров [52, 54, 94, 129, 556].