
Поступление СОг в легких из крови в альвеолы обеспечивается из следующих источников: 1) из СОг, растворенного в плазме крови (5—10%); 2) из гидрокарбонатов (80—90%); 3) из карбаминовых соединений эритроцитов (5—15%), которые способны диссоциировать.
‘ Для СОг коэффициент растворимости в мембранах аэрогемати- ческого барьера больше, чем для Ог, и составляет в среднем 0,231 ммольжл-1кПа-1, поэтому СОг диффундирует быстрее, чем Ог. Это положение является верным только для диффузии молекулярного СОг. Большая часть СОг транспортируется в организме в связанном состоянии в виде гидрокарбонатов и карбаминовых соединений, что увеличивает время обмена СОг, затрачиваемое на диссоциацию этих соединений.
В венозной крови, притекающей к капиллярам легких, напряжение СОг составляет в среднем 46 мм рт. ст. (6,1 кПа), а в альвеолярном воздухе парциальное давление СОг равно в среднем 40 мм рт. ст. (5,3 кПа), что обеспечивает диффузию СОг из плазмы крови в альвеолы легких по концентрационному градиенту.
Эндотелий капилляров проницаем только для молекулярного СОг как полярной молекулы (О — С — О). Из крови в альвеолы диффундирует физически растворенный в плазме крови молекулярный СОг. Кроме того, в альвеолы легких диффундирует СОг, который высвобождается из карбаминовых соединений эритроцитов благодаря реакции окисления гемоглобина в капиллярах легкого, а также из гидрокарбонатов плазмы крови в результате их быстрой диссоциации с помощью фермента карбоангидразы, содержащейся в эритроцитах.
Молекулярный СОг проходит аэрогематический барьер, а затем поступает в альвеолы.
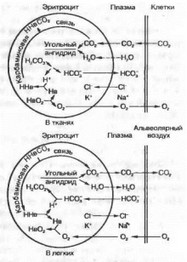
Рис. 8.8. Участие эритроцитов в обмене Ог и СОг в тканях и в легких.
 В норме через 1 с происходит выравнивание концентраций СОг на альвеолярно-капиллярной мембране, поэтому за половину времени капиллярного кровотока происходит полный обмен СОг через аэрогематический барьер. Реально равновесие наступает несколько медленнее. Это связано с тем, что перенос СОг, так же как и Ог, ограничивается скоростью перфузии капилляров легких.
В норме через 1 с происходит выравнивание концентраций СОг на альвеолярно-капиллярной мембране, поэтому за половину времени капиллярного кровотока происходит полный обмен СОг через аэрогематический барьер. Реально равновесие наступает несколько медленнее. Это связано с тем, что перенос СОг, так же как и Ог, ограничивается скоростью перфузии капилляров легких.
Диффузия СО2 из тканей в кровь. Обмен СОг между клетками тканей с кровью тканевых капилляров осуществляется с помощью следующих реакций: 1) обмена С1-и НСОГ через мембрану эритроцита; 2) образования угольной кислоты из гидрокарбонатов;
- диссоциации угольной кислоты и гидрокарбонатов (рис. 8.8).
В ходе газообмена СОг между тканями и кровью содержание НСОГ в эритроците повышается и они начинают диффундировать в кровь. Для поддержания электронейтральности в эритроциты начнут поступать из плазмы дополнительно ионы С1Т Наибольшее количество бикарбонатов плазмы крови образуется при участии карбоангидразы эритроцитов.
Карбаминовый комплекс СОг с гемоглобином образуется в результате реакции СОг с радикалом NH7 глобина. Эта реакция протекает без участия какого-либо фермента, т. е. она не нуждается в катализе. Реакция СОг с НЬ приводит, во-первых, к высвобождению Н+; во-вторых, в ходе образования карбаминовых комплексов снижается сродство НЬ к Ог. Эффект сходен с действием низкого pH. Как известно, в тканях низкое pH потенцирует высвобождение Ог из оксигемоглобина при высокой концентрации СОг (эффект Бора).
С другой стороны, связывание 02 гемоглобином снижает сродство его аминогрупп к COz (эффект Холдена).
Каждая реакция в настоящее время хорошо изучена. Например, полупериод обмена С1-и НСОГ равен 0,11—0,16 с при 37 °С. В условиях in vitro образование молекулярного COz из гидрокарбонатов происходит чрезвычайно медленно и диффузия этого газа занимает около 5 мин, тогда как в капиллярах легкого равновесие наступает через 1 с. Это определяется функцией фермента карбоангидразы угольной кислоты. В функции карбоангидразы выделяют следующие типы реакций:
С02 +Н20 X Н2СОэ X Н+ +НСОэ.
Процесс выведения С02 из крови в альвеолы легкого менее лимитирован, чем оксигенация крови. Это обусловлено тем, что молекулярный С02 легче проникает через биологические мембраны, чем 02. По этой причине он легко проникает из тканей в кровь. К тому же карбоангидраза способствует образованию гидрокарбоната. Яды, которые ограничивают транспорт 02 (такие как СО, метгемоглобинобразующие субстанции — нитриты, метиленовый синий, ферроцианиды и др.) не действуют на транспорт С02. Блокаторы карбоангидразы, например диакарб, которые используются нередко в клинической практике или для профилактики горной или высотной болезни, полностью никогда не нарушают образование молекулярного С02. Наконец, ткани обладают большой буферной емкостью, но не защищены от дефицита 02. По этой причине нарушение транспорта 02 наступает в организме гораздо чаще и быстрее, чем нарушения газообмена С02. Тем не менее при некоторых заболеваниях высокое содержание С02 и ацидоз могут быть причиной смерти.
Измерение напряжения 02 и С02 в артериальной или смешанной венозной крови производят полярографическими методами с использованием очень небольшого количества крови. Количество газов в крови измеряют после их полного извлечения из пробы крови, взятой для анализа.
Такие исследования выполняют с помощью манометрических приборов типа аппарата Ван-Слайка, или гемоалкариметра (необходимо 0,5—2,0 мл крови) или на микроманометре Холандера (необходимо около 50 мкл крови).