
Нервная экстракардиальная регуляция. Эта регуляция осуществляется импульсами, поступающими к сердцу из ЦНС по блуждающим и симпатическим нервам.
Подобно всем вегетативным нервам, сердечные нервы образованы двумя нейронами. Тела первых нейронов, отростки которых составляют блуждающие нервы (парасимпатический отдел автономной нервной системы), расположены в продолговатом мозге (рис. 7.11). Отростки этих нейронов заканчиваются в интрамуральных ганглиях сердца. Здесь находятся вторые нейроны, отростки которых идут к проводящей системе, миокарду и коронарным сосудам.
Первые нейроны симпатической части автономной нервной системы, передающие импульсы к сердцу, расположены в боковых рогах пяти верхних сегментов грудного отдела спинного мозга. Отростки этих нейронов заканчиваются в шейных и верхних грудных симпатических узлах. В этих узлах находятся вторые нейроны, отростки которых идут к сердцу. Большая часть симпатических нервных волокон, иннервирующих сердце, отходит от звездчатого узла.
Влияние на сердце блуждающих нервов впервые изучили братья Вебер (1845). Они установили, что раздражение этих нервов тормозит работу сердца вплоть до полной его остановки в диастолу. Это был первый случай обнаружения в организме тормозящего влияния нервов.
При электрическом раздражении периферического отрезка перерезанного блуждающего нерва происходит урежение сердечных сокращений. Это явление называется отрицательным хронотропным эффектом. Одновременно отмечается уменьшение амплитуды сокращений — отрицательный инотропный эффект.
При сильном раздражении блуждающих нервов работа сердца на некоторое время прекращается. В этот период возбудимость
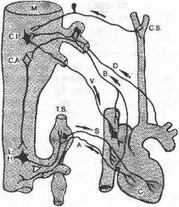 Рис. 7.11. Экстракардиальиые нервы сердца.
Рис. 7.11. Экстракардиальиые нервы сердца.
С — сердце; М — продолговатый мозг; С. 1. — ядро, вызывающее торможение деятельности сердца; С. А. — ядро, вызывающее учашение сердечной деятельности; L. Н. — боковой рог спинного мозга; Т. S. — симпатический ствол; V — эфферентные волокна блуждающего нерва; D — нерв «депрессор» (афферентные волокна блуждающего нерва); S — симпатические волокна; А — спинномозговые афферентные волокна; С. S. — каротидный синус; В — афферентные волокна от правого предсердия и полой вены.
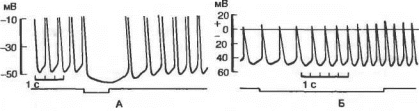
Рис. 7.12. Потенциалы действия клетки — водителя ритма сердца лягушки при раздражении блуждающего (А) и симпатического (Б) нервов.
На рисунке Л показана только нижняя половина потенциала действия.
мышцы сердца понижена. Понижение возбудимости мышцы сердца называется отрицательным батмотропным эффектом. Замедление проведения возбуждения в сердце называется отрицательным дромотропным эффектом. Нередко наблюдается полная блокада проведения возбуждения в предсердно-желудочковом узле.
Микроэлектродные отведения потенциалов от одиночных мышечных волокон предсердий показали увеличение мембранного потенциала — гиперполяризацию при сильном раздражении блуждающего нерва (рис. 7.12).
При продолжительном раздражении блуждающего нерва прекратившиеся вначале сокращения сердца восстанавливаются, несмотря на продолжающееся раздражение. Это явление называют ускользанием сердца из-под влияния блуждающего нерва.
Влитие на сердце симпатических нервов впервые было изучено братьями Цион (1867), а затем И. П. Павловым. Ционы описали учащение сердечной деятельности при раздражении симпатических нервов сердца (положительный хронотропный эффект); соответствующие волокна они назвали nn. accelerantes cordis (ускорители сердца).
При раздражении симпатических нервов ускоряется спонтанная деполяризация клеток — водителей ритма в диастолу, что ведет к учащению сердечных сокращений.
Раздражение сердечных ветвей симпатического нерва улучшает проведение возбуждения в сердце (положительный дромотропный эффект) и повышает возбудимость сердца (положительный бат- мотропный эффект). Влияние раздражения симпатического нерва наблюдается после большого латентного периода (10 с и более) и продолжается еще долго после прекращения раздражения нерва.
И. П. Павлов (1887) обнаружил нервные волокна (усиливающий нерв), усиливающие сердечные сокращения без заметного учащения ритма (положительный инотропный эффект).
Инотропный эффект «усиливающего» нерва хорошо виден при регистрации внутрижелудочкового давления электроманометром. Выраженное влияние «усиливающего» нерва на сократимость миокарда проявляется особенно при нарушениях сократимости. Одной из таких крайних форм нарушения сократимости является альтер-
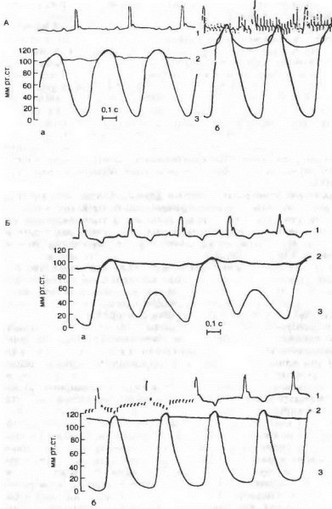
Рис. 7.13. Влияние «усиливающего» нерва на сердце (по В. М. Покровскому).
Л — влияние «усиливающего нерва* на динамику сокращений сердив; Б — устранение «усиливающим* нервом альтернации силы сокращений сердив; а — до раздражения, б — во время раздражения нерва. I — ЭКГ; 2 — давление в аорте; 3 — давление в левом желудочке до раздражения и во время раздражения нерва.
нация сердечных сокращений, когда одно «нормальное» сокращение миокарда (в желудочке развивается давление, превышающее давление в аорте и осуществляется выброс крови из желудочка в аорту) чередуется со «слабым» сокращением миокарда, при котором давление в желудочке в систолу не достигает давления в аорте и выброса крови не происходит. «Усиливающий» нерв не только усиливает обычные сокращения желудочков, но и устраняет альтернацию, восстанавливая неэффективные сокращения до обычных (рис. 7.13). По мнению И. П. Павлова, эти волокна являются специально трофическими, т. е. стимулирующими процессы обмена веществ.
Совокупность приведенных данных позволяет представить влияние нервной системы на ритм сердца как корригирующее, т. е. ритм сердца зарождается в его водителе ритма, а нервные влияния ускоряют или замедляют скорость спонтанной деполяризации клеток водителя ритма, ускоряя или замедляя таким образом частоту сердцебиений.
В последние годы стали известны факты, свидетельствующие о возможности не только корригирующих, но и пусковых влияний нервной системы на ритм сердца, когда сигналы, приходящие по нервам, инициируют сокращения сердца. Это можно наблюдать в опытах с раздражением блуждающего нерва в режиме, близком к естественной импульсации в нем, т. е. «залпами» («пачками») импульсов, а не непрерывным потоком, как это делалось традиционно. При раздражении блуждающего нерва «залпами» импульсов сердце сокращается в ритме этих «залпов» (каждому «залпу» соответствует одно сокращение сердца). Меняя частоту и характеристику «залпов», можно управлять ритмом сердца в широких пределах.
Воспроизведение сердцем центрального ритма резко изменяет электрофизиологические параметры деятельности синоатриального узла. При работе узла в режиме автоматии, а также при изменениях частоты под влиянием раздражения блуждающего нерва в традиционном режиме возбуждение возникает в одной точке узла, в случае воспроизведения центрального ритма в инициации возбуждения принимает участие одновременно множество клеток узла. На изохронной карте движения возбуждения в узле этот процесс отражается не в виде точки, а в виде большой площади, образованной одновременно возбуждающимися структурными элементами. Сигналы, обеспечивающие синхронное воспроизведение сердцем центрального ритма, отличаются по своей медиаторной природе от общетормозных влияний блуждающего нерва. По-видимому, выделяющиеся в этом случае наряду с ацтилхолином регуляторные пептиды отличаются по своему составу, т. е. реализация каждого типа эффектов блуждающего нерва обеспечивается своей смесью медиаторов («ме- диаторные коктейли»).
С целью изменения частоты посылки «пачек» импульсов из сердечного центра продолговатого мозга у людей можно воспользоваться такой моделью. Человеку предлагают дышать чаще, чем сокращается его сердце. Для этого он следит за миганием лампочки фотостимулятора и на каждую вспышку света производит одно

дыхание. Фотостимулятор устанавливается с частотой, превышающей исходную частоту сердцебиений. За счет иррадиации возбуждения с дыхательных на сердечные нейроны в продолговатом мозге в сердечных эфферентных нейронах блуждающего нерва формируются «пачки» импульсов в новом, общем для дыхательных и сердечных центров, ритме. При этом синхронизация ритмов дыхания и сердцебиения достигается за счет «залпов» импульсов, приходящих к сердцу по блуждающим нервам. В опытах на собаках фенбмен синхронизации дыхательных и сердечных ритмов наблюдается при резком учащении дыхания во время перегревания. Как только ритм учащающегося дыхания станет равным частоте сердцебиений, оба ритма синхронизируются и учащаются или урежаются в определен
ном диапазоне синхронно. Если при этом нарушить проведение сигналов по блуждающим нервам посредством их перерезки или холодовой блокады, то синхронизация ритмов исчезнет. Следовательно, и в этой модели сердце сокращается под влиянием «залпов» импульсов, приходящих к нему по блуждающим нервам.
Совокупность изложенных экспериментальных фактов позволила сформировать представление о существовании наряду с внут- рисердечным и центрального генератора ритма сердца (В. М. Покровский). При этом последний в естественных условиях формирует адаптивные (приспособительные) реакции сердца, воспроизводя ритм сигналов, приходящих к сердцу по блуждающим нервам. Внутрисердечный генератор обеспечивает поддержание жизни за счет сохранения насосной функции сердца в случае выключения центрального генератора при наркозе, ряде заболеваний, обмороке и т. д.
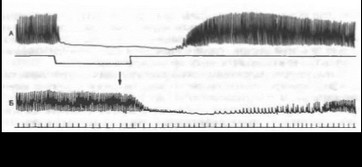
Химический механизм передачи нервных импульсов в сердце. При раздражении периферических отрезков блуждающих нервов в их окончаниях в сердце выделяется АХ, а при раздражении симпатических нервов — норадреналин. Эти вещества являются непосредственными агентами, вызывающими торможение или усиление деятельности сердца, и поэтому получили название медиаторов (передатчиков) нервных влияний. Существование медиаторов было показано Леви (1921). Он раздражал блуждающий или симпатический нерв изолированного сердца лягушки, а затем переносил жидкость из этого сердца в другое, тоже изолированное, но не подвергавшееся нервному влиянию — второе сердце давало такую же реакцию (рис. 7.14, 7.15). Следовательно, при раздражении нервов первого сердца в питающую его жидкость переходит соответствующий медиатор. На нижних кривых можно видеть эффекты, вызываемые перенесенным раствором Рингера, находившимся в сердце во время раздражения.
АХ, образующийся в окончаниях блуждающего нерва, быстро разрушается ферментом холинэстеразой, присутствующим в крови и клетках, поэтому АХ оказывает только местное действие. Норадреналин разрушается значительно медленнее, чем АХ, и потому действует дольше. Этим объясняется то, что после прекращения раздражения симпатического нерва в течение некоторого времени сохраняются учащение и усиление сердечных сокращений.
Получены данные, свидетельствующие о том, что при возбуждении наряду с основным медиаторным веществом в синаптическую щель поступают и другие биологически активные вещества, в частности пептиды. Последние обладают модулирующим действием, изменяя величину и направленность реакции сердца на основной медйатор. Так, опиоцдные пептиды угнетают эффекты раздражения блуждающего нерва, а пептид дельта-сна усиливает вагусную брадикардию.