Секреторная функция желудка
Образование, состав и свойства желудочного сока. Желудочный сок продуцируется железами желудка, расположенными в его слизистой оболочке. Она покрыта слоем цилиндрического эпителия, клетки которого секретируют слизь и слабощелочную жидкость. Слизь секретируется в виде густого геля, который покрывает равномерным слоем всю слизистую оболочку.
На поверхности слизистой оболочки видны мелкие впадинки — желудочные ямки. Общее их количество достигает 3 млн. В каждую из них открываются просветы 3—7 трубчатых желудочных желез. Различают три вида желудочных желез: собственные железы желудка, кардиальные и пилорические.
Собственные железы желудка располагаются в области тела и дна желудка (фундальные). Фундальные железы состоят из трех основных типов клеток: главные клетки — секретирующие пепси- ногены, обкладочные (париетальные, оксинтные гландулоциты) — соляную кислоту и добавочные — слизь. Соотношение разных типов клеток в железах слизистой оболочки различных отделов желудка неодинаково. Кардиальные железы расположены в кардиальном отделе желудка — это трубчатые железы, состоящие в основном из клеток, продуцирующих слизь. В пилорическом отделе железы практически не имеют обкладочных клеток. Пилорические железы выделяют небольшое количество секрета, нестимулируе- мое приемом пищи. Ведущее значение в желудочном пищеварении имеет желудочный сок, вырабатываемый фундальными железами.
За сутки желудок человека выделяет 2—2,5 л желудочного сока. Он представляет собой бесцветную прозрачную жидкость, содержащую соляную кислоту (0,3—0,5%) и поэтому имеющую кислую реакцию (pH 1,5—1,8). Величина pH содержимого желудка значительно выше, так как сок фундальных желез частично нейтрализуется принятой пищей.
В желудочном соке имеются многие неорганические вещества: вода (995 г/л), хлориды (5—6 г/л), сульфаты (10 мг/л), фосфаты (10—60 мг/л), гидрокарбонаты (0—1,2 г/л) натрия, калия, кальция, магния, аммиак (20—80 кг/л). Осмотическое давление желудочного сока выше, чем плазмы крови.
Обкладочные клетки продуцируют соляную кислоту одинаковой концентрации (160 ммоль/л), но кислотность выделяющегося сока варьирует за счет изменения числа функционирующих париетальных гландулоцитов и нейтрализации соляной кислоты щелочными компонентами желудочного сока. Чем быстрее секреция соляной кислоты, тем меньше она нейтрализуется и тем выше кислотность желудочного сока.
Синтез соляной кислоты в обкладочных клетках сопряжен с клеточным дыханием и является аэробным процессом; при гипоксии секреция кислоты прекращается. Согласно «карбоангидраз- ной» гипотезе, ионы Н+ для синтеза соляной кислоты получаются в результате гидратации СОг и диссоциации образовавшейся при этом Н2СО3. Этот процесс катализируется ферментом карбоан- гидразой. Согласно «редокс»-гипотезе, ионы Н+ для синтеза соляной кислоты поставляются митохондриальной дыхательной цепью, а транспорт ионов Н+и С1 осуществляется за счет энергии окислительно-восстановительных цепей. «АТФазная» гипотеза утверждает, что для транспорта этих ионов используется энергия АТФ, а Н+ могут происходить из различных источников, в том числе поставляться карбоангидразой из фосфатной буферной системы.
Сложные процессы, завершающиеся синтезом и экструзией из обкладочных клеток соляной кислоты, включают в себя три звена: 1) реакции фосфорилирования—дефосфорилирования; 2) митохондриальную окислительную цепь, работающую в режиме помпы; т. е. переносящую протоны из матриксного пространства вовне;
- Н+, К+-АТФазу секреторной мембраны, осуществляющую «перекачку» этих протонов из клетки в просвет желез за счет энергии АТФ.
Соляная кислота желудочного сока вызывает денатурацию и набухание белков и тем самым способствует их последующему расщеплению пепсинами, активирует пепсиногены, создает кислую среду, необходимую для расщепления пищевых белков пепсинами; участвует в антибактериальном действии желудочного сока и регуляции деятельности пищеварительного тракта (в зависимости от pH его содержимого усиливается или тормозится нервными механизмами и гастроинтестинальными гормонами его деятельность).
Органические компоненты желудочного сока представлены азотсодержащими веществами (200—500 мг/л): мочевиной, мочевой и молочной кислотами, полипептидами. Содержание белка достигает 3 г/л, мукопротеидов — до 0,8 г/л, мукопротеаз — до 7 г/л. Органические вещества желудочного сока являются продуктами секреторной деятельности желудочных желез и обмена веществ в слизистой оболочке желудка, а также транспортируются через нее из крови. В числе белков особое значение для пищеварения имеют ферменты.
Главные клетки желудочных желез синтезируют несколько пепсиногенов, которые принято делить на две группы. Пепсиногены первой группы локализуются в фундальной части желудка, второй группы — в антральной части и начале двенадцатиперстной кишки. При активации пепсиногенов путем отщепления от них полипептида образуется несколько пепсинов. Собственно пепсинами принято называть ферменты класса протеаз, гидролизующие белки с максимальной скоростью при pH 1,5—2,0. Протеаза, названная гастриксином, имеет оптимальный для гидролиза белков pH 3,2—
- Соотношение содержания пепсина и гастриксина в желудочном соке человека колеблется от 1:2 до 1:5. Эти ферменты различаются действием на разные виды белков.
Пепсины являются эндопептидазами, и основными продуктами их гидролитического действия на белки являются полипептиды (разрываются около 10% связей с освобождением аминокислот). Способность пепсинов гидролизовывать белки в широком диапазоне pH имеет большое значение для желудочного протеолиза, который происходит при разном pH в зависимости от объема и кислотности желудочного сока, буферных свойств и количества принятой пищи, диффузии кислого сока в глубь пищевого желудочного содержимого. Гидролиз белков происходит в непосредственной близости от слизистой оболочки. Проходящая перистальтическая волна «снимает» («слизывает») примукозальный слой, продвигает его к антральной части желудка, в результате чего к слизистой оболочке примыкает бывший более глубокий слой пищевого содержимого, на белки которого пепсины действовали при слабокислой реакции. Эти белки подвергаются гидролизу пепсинами в более кислой среде.
Важным компонентом желудочного сока являются мукоиды, продуцируемые мукоцитами поверхностного эпителия, шейки фун- дальных и пилорических желез (до 15 г/л). К мукоидам относится и гастромукопротеид (внутренний фактор Касла). Слой слизи толщиной 1—1,5 мм защищает слизистую оболочку желудка и называется слизистым защитным барьером желудка. Слизь — мукоидный секрет — представлена в основном двумя типами веществ — гликопротеинами и протеогликанами.
Сок, выделяемый разными участками слизистой оболочки желудка, содержит различное количество пепсиногена и соляной кислоты. Так, железы малой кривизны желудка продуцируют сок с более высокими кислотностью и содержанием пепсина, чем железы большой кривизны желудка.
Железы в пилорической части желудка выделяют небольшое количество сока слабощелочной реакции с большим содержанием слизи. Увеличение секреции происходит при местном механическом и химическом раздражении пилорической части желудка. Секрет пилорических желез обладает небольшой протеолитической, липолитической и амилолитической активностью. Существенного значения в желудочном пищеварении ферменты, обусловливающие эту активность, не имеют. Щелочной пилорический секрет частично нейтрализует кислое содержимое желудка, эвакуируемое в двенадцатиперстную кишку.
Показатели желудочной секреции имеют существенные индивидуальные, половые и возрастные различия. При патологии желудочная секреция может повышаться (гиперсекреция) или понижаться (гипосекреция), соответственно может меняться секреция соляной кислоты (гипер- и гипоацидность, отсутствие ее в соке — анацидность, ахлоргидрия). Меняется содержание пепсиногенов и соотношение их видов в желудочном соке.
Большое защитное значение имеет слизистый барьер желудка, разрушение которого может быть одной из причин повреждения слизистой оболочки желудка и даже глубже расположенных структур его стенки. Этот барьер повреждается при высокой концентрации в содержимом желудка соляной кислоты, алифатическими кислотами (уксусная, соляная, масляная, про- пионовая) даже в небольшой концентрации, детергентами (желчные кислоты, салициловая и сульфосалициловая кислоты в кислой среде желудка), фосфолипазами, алкоголем. Длительный контакт этих веществ (при их относительно высокой концентрации) нарушает слизистый барьер и может привести к повреждению сли-
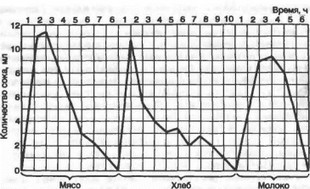
Рис. 9.11. Кривые сокоотделения павловского желудочка на мясо, хлеб и молоко.
зистой оболочки желудка. Разрушению слизистого барьера и стимуляции секреции соляной кислоты способствует деятельность микроорганизмов Helicobacter pylori. В кислой среде и в условиях нарушенного слизистого ьарьера возможно переваривание элементов слизистой оболочки пепсином (пептический фактор язвообра- зования). Этому способствует также снижение секреции гидрокарбонатов и микроциркуляции крови в слизистой оболочке желудка.
Регуляция желудочной секреции. Вне пищеварения железы желудка выделяют небольшое количество желудочного сока. Прием пищи резко увеличивает его выделение. Это происходит за счет стимуляции желудочных желез нервными и гуморальными механизмами, составляющими единую систему рефляции. Стимулирующие и тормозные регуляторные факторы обеспечивают зависимость сокоотделения желудка от вида принимаемой пищи. Эта зависимость была впервые обнаружена в лаборатории И. П. Павлова в опытах на собаках с изолированным павловским желудочком, которым скармливалась различная пища. Объем и характер секреции во времени, кислотность и содержание в соке пепсинов определяются видом принятой пищи (рис. 9.11).
Стимуляция секреции соляной кислоты обкладочными клетками осуществляется непосредственно и опосредованно через другие механизмы. Непосредственно стимулируют секрецию соляной кислоты обкладочными клетками холинергические волокна блуждающих нервов, медиатор которых — ацетилхолин (АХ) — возбуждает М-холинорецепторы базолатеральных мембран гландулоцитов. Эффекты АХ и его аналогов блокируются атропином. Непрямая стимуляция клеток блуждающими нервами опосредуется также гастрином и гистамином.
Гастрин высвобождается из G-клеток, основное количество которых находится в слизистой оболочке пилорической части желудка. После хирургического удаления пилорической части желу-
дочная секреция резко снижается. Высвобождение гастрина усиливается импульсами блуждающего нерва, а также местным механическим и химическим раздражением этой части желудка. Химическими стимуляторами G-клеток являются продукты переваривания белков — пептиды и некоторые аминокислоты, экстрактивные вещества мяса и овощей. Бели pH в антральной части желудка понижается, что обусловлено повышением секреции соляной кислоты железами желудка, то высвобождение гастрина уменьшается, а при pH 1,0 прекращается и объем секреции резко понижается. Таким образом, гастрин принимает участие в саморегуляции желудочной секреции в зависимости от величины pH содержимого антрального отдела. Гастрин в наибольшей мере стимулирует париетальные гландулоциты желудочных желез и увеличивает выделение соляной кислоты.
К стимуляторам обкладочных клеток желудочных желез относится и гистамин, образующийся в ECL-клетках слизистой оболочки желудка. Высвобождение гистамина обеспечивается гастри- ном. Гистамин стимулирует гландулоциты, влияя на Нг-рецепторы их мембран и вызывая выделение большого количества сока высокой кислотности, но бедного пепсином.
Стимулирующие эффекты гастрина и гистамина зависят от сохранности иннервации желудочных желез блуждающими нервами: после хирургической и фармакологической ваготомии секреторные эффекты этих гуморальных стимуляторов понижаются.
Желудочную секрецию возбуждают также всосавшиеся в кровь продукты переваривания белков.
Торможение секреции соляной кислоты вызывают секретин, ХЦК, глюкагон, ЖИП, ВИП, нейротензин, полипептид УУ, сома- тостатин, тиролиберин, энтерогастрон, АДГ, кальцитонин, окси- тоцин, простагландин ПГЕ2, бульбогастрон, кологастрон, серотонин (см. табл. 9.2). Высвобождение некоторых из них в соответствующих эндокринных клетках слизистой оболочки кишечника контролируется свойствами химуса. В частности, торможение желудочной секреции жирной пищей в большой мере обусловлено влиянием на железы желудка ХЦК. Повышение кислотности содержимого двенадцатиперстной кишки тормозит выделение соляной кислоты железами желудка. Торможение секреции осуществляется рефлекторно, а также вследствие образования гормонов двенадцатиперстной кишки.
Механизм стимуляции и торможения секреции соляной кислоты различными нейротрансмиттерами и гормонами неодинаков. Так, АХ усиливает секрецию кислоты обкладочными клетками путем активации мембранной Na+, К+-АТФазы, увеличения транспорта ионов Са?+ и эффектов повышенного внутриклеточного содержания цГМФ, высвобождения гастрина и потенцирования его влияния.
Гастрин усиливает секрецию соляной кислоты посредством гистамина, а также путем действия на мембранные рецепторы гастрина и усиления внутриклеточного транспорта ионов Са2+. Гистамин стимулирует секрецию обкладочных клеток через их мембранные Н2-рецепторы и систему аденилатциклаза (АЦ) — цАМФ.
Стимуляторами секреции пепсиногена главными клетками являются холинергические волокна блуждающих нервов, гастрин, гистамин, симпатические волокна, оканчивающиеся на р-адрено- рецепторах, секретин и ХЦК. Усиление секреции пепсиногенов главными клетками желудочных желез осуществляется несколькими механизмами. Среди них увеличение переноса ионов Са?+ в клетку и стимуляция Na+, К+-АТФазы; усиление внутриклеточного перемещения гранул зимогена, активация мембранной фосфорилазы, что усиливает их прохождение через апикальные мембраны, активация системы цГМФ и цАМФ.
Эти механизмы в неодинаковой мере активируются или тормозятся различными нейротрансмиттерами и гормонами, непосредственными и опосредованными влияниями их на главные клетки и секрецию пепсиногена. Показано, что гистамин и гастрин влияют на него опосредованно — усиливают секрецию соляной кислоты, а снижение pH содержимого желудка через местный холинергический рефлекс усиливает секрецию главных клеток. Описано и прямое стимулирующее влияние на них гастрина. В высоких дозах гистамин тормозит их секрецию. ХЦК, секретин и р-адреномиме- тики непосредственно стимулируют секрецию главных клеток, но тормозят секрецию обкладочных, что свидетельствует о существовании на них разных рецепторов регуляторных пептидов.
Стимуляция секреции слизи мукоцитами осуществляется холинергическими волокнами блуждающих нервов. Гастрин и гистамин умеренно стимулируют мукоциты, видимо, в связи с удалением слизи с их мембран при выраженной секреции кислого желудочного сока. Ряд ингибиторов секреции соляной кислоты — серотонин, соматостатин, адреналин, дофамин, энкефалин, простаглан- дин ПГЕ2 — усиливает секрецию слизи. Полагают, что ПГЕ2 усиливает секрецию слизи названными веществами.
При приеме пищи и пищеварении в усиленно секретирующих железах желудка кровоток возрастает, что обеспечивается действием холинергических нервных механизмов, пептидов пищеварительного тракта и местных вазодилататоров. В слизистой оболочке кровоток нарастает интенсивнее, чем в подслизистой основе и мышечном слое желудочной стенки.
Фазы желудочной секреции. Нервные, гуморальные факторы и паракринные механизмы тонко регулируют секрецию желез желудка, обеспечивают выделение определенного количества сока, кислото- и ферментовыделение в зависимости от количества и качества принятой пищи, эффективности ее переваривания в желудке и тонкой кишке. Происходящую при этом секрецию принято делить на три фазы.
Начальная секреция желудка возникает рефлекторно в ответ на раздражение дистантных рецепторов, возбуждаемых видом и запахом пищи, всей обстановкой, связанной с ее приемом (условнорефлекторные раздражения). Кроме того, секреция желудка возбуждается рефлекторно в ответ на раздражение принимаемой пищей рецепторов полости рта и глотки (безусловнорефлекторные раздражения). Эти рефлексы обеспечивают пусковые влияния на железы желудка. Желудочную секрецию, обусловленную этими сложными рефлекторными влияниями, принято называть первой, или мозговой, фазой секреции (см. рис. 9.8).
Механизмы первой фазы секреции желудка были изучены в опытах на эзофаготомированных собаках с фистулой желудка. При кормлении такой собаки пища выпадает из пищевода и не поступает в желудок, однако через 5—10 мин после начала мнимого кормления начинает выделяться желудочный сок. Аналогичные данные были получены при исследовании людей, страдающих сужением пищевода и подвергшихся вследствие этого операции наложения фистулы желудка. Жевание пищи вызывало у людей выделение желудочного сока.
Рефлекторные влияния на желудочные железы передаются через блуждающие нервы. После их перерезки у эзофаготомирован- ной собаки ни мнимое кормление, ни вид и запах пищи не вызывают секреции. Если раздражать периферические концы перерезанных блуждающих нервов, то отмечается выделение желудочного сока с высоким содержанием в нем соляной кислоты и пепсина.
В стимуляцию желудочных желез в первую фазу включен и гастриновый механизм. Доказательством этого служит увеличение содержания гастрина в крови людей при мнимом кормлении. После удаления пилорической части желудка, где продуцируется гас- трин, секреция в первую фазу понижается.
Секреция в мозговую фазу зависит от возбудимости пищевого центра и может легко тормозиться при раздражении различных внешних и внутренних рецепторов. Так, плохая сервировка стола, неопрятность места приема пищи снижают и тормозят желудочную секрецию. Оптимальные условия приема пищи положительно влияют на желудочную секрецию. Прием в начале еды сильных пищевых раздражителей повышает желудочную секрецию в первую фазу.
На секрецию первой фазы наслаивается секреция второй фазы, которая называется желудочной, так как обусловлена влиянием пищевого содержимого в период его нахождения в желудке. Наличие этой фазы секреции доказывается тем, что вкладывание пищи в желудок через фистулу, вливание через нее или зонд растворов в желудок, раздражение его механорецепторов вызывают отделение желудочного сока. Объем секреции при этом в 2—3 раза меньше, чем при естественном приеме пищи. Это подчеркивает большое значение пусковых рефлекторных влияний, осуществляемых преимущественно в первую фазу на желудочные железы. Во вторую фазу железы желудка испытывают в основном корригирующие влияния. Эти влияния путем усиления и ослабления деятельности желез обеспечивают соответствие секреции количеству и свойствам пищевого желудочного содержимого, т. е. осуществляют коррекцию секреторной деятельности желудка.
Сокоотделение при механическом раздражении желудка возбуждается рефлекторно с механорецепторов слизистой оболочки и мышечного слоя стенки желудка. Секреция резко уменьшается после перерезки блуждающих нервов. Кроме того, механическое раздражение желудка, особенно его пилорической части, приводит к высвобождению из G-клеток гастрина.
Повышение кислотности содержимого антральной части желудка тормозит высвобождение гастрина и снижает желудочную секрецию. В фундальной части желудка кислотность его содержимого рефлекторно усиливает секрецию, особенно выделение пепсиноге- на. Определенное значение в реализации желудочной фазы секреции имеет гистамин, значительное количество которого образуется в слизистой оболочке желудка.
Мясной бульон, капустный сок, продукты гидролиза белков при введении в тонкую кишку вызывают выделение желудочного сока. Нервные влияния с рецепторов кишечника на железы желудка обеспечивают секрецию в третью, кишечную, фазу. Возбуждающие и тормозные влияния из двенадцатиперстной и тощей кишки на железы желудка осуществляются с помощью нервных и гуморальных механизмов, корригирующих секрецию. Нервные влияния передаются с механо- и хеморецепторов кишечника. Стимуляция желудочных желез в кишечную фазу является прежде всего результатом поступления в двенадцатиперстную кишку недостаточно физически и химически обработанного содержимого желудка. В стимуляции желудочной секреции принимают участие всосавшиеся в кровь продукты гидролиза питательных веществ, особенно белков. Эти вещества могут возбуждать железы желудка опосредованно через гастрин и гистамин, а также непосредственно действуя на желудочные железы.
Торможение желудочной секреции в ее кишечную фазу вызы- вается рядом веществ в составе кишечного содержимого, которые по убывающей силе тормозного действия расположены в следующем порядке: продукты гидролиза жира, полипептиды, аминокислоты, продукты гидролиза крахмала, Н+ (pH ниже 3 оказывает сильное тормозное действие).
Высвобождение в двенадцатиперстной кишке секретина и ХЦК под влиянием поступившего в кишечник содержимого желудка и образовавшихся продуктов гидролиза питательных веществ тормозит секрецию соляной кислОты, но усиливает секрецию пепсино- гена. Желудочную секрецию тормозят и другие кишечные гормоны из группы гастронов и глюкагой, а также серотонин.
Влияние пищевых режимов на желудочную секрецию. В экспериментах на животных И. П. Павловым с сотрудниками, а затем И. П. Разенковым с сотрудниками показано, что секреция желудочных желез значительно изменяется в зависимости от характера питания. При длительном (30—40 дней) употреблении пищи, содержащей большое количество углеводов (хлеб, овощи), секреция уменьшается (в основном во вторую и третью фазы). Если животное длительный срок (30—60 дней) принимает пищу, богатую белками, например мясо, то секреция увеличивается, в особенности во вторую и третью фазы. При этом меняются не только объем и динамика во времени желудочной секреции, но и ферментативные свойства желудочного сока. А. М. Уголевым экспериментально установлено, что длительный прием растительной пищи повышает активность желудочного сока по отношению к белкам растительного происхождения («фитолитическая активность»), а преобладание в пищевом рационе животных белков повышает способность желудочного сока гидролизовать их («зооли- тическая активность»). Это связано с изменением кислотности сока и соотношения в нем видов и свойств пепсинов.
А так же в разделе «Секреторная функция желудка »
- Прием пищи
- Жевание
- Слюноотделение
- Глотание
- ПИЩЕВАРЕНИЕ В ЖЕЛУДКЕ
- Моторная функция желудка
- Эвакуация содержимого желудка в двенадцатиперстную кишку
- Рвота
- ПИЩЕВАРЕНИЕ В ТОНКОЙ КИШКЕ
- Секреция поджелудочной железы
- Желчеотделение и желчевыделение
- Кишечная секреция
- Полостное и пристеночное пищеварение в тонкой кишке
- Моторная функция тонкой кишки
- Всасывание различных веществ в тонкой кишке