I. ОСНОВНЫЕ ПОНЯТИЯ
А. Антиген и антитело
- Антиген (Ar) — вещество, несущее признаки генетически чужеродной информации. Ar можно определить как молекулу, распознаваемую иммунокомпетентными клетками как чужеродную (не свою). Иммуногены — Ar, вызывающие в организме иммунный ответ. Молекула иммуногена взаимодействует с AT или рецептором Т-лимфоцитов [глава 6.1 II Б 6 е].
а. Экзогенные иммуногены подвергаются эндоцитозу и расщеплению в Аг-представ- ляющих клетках (I В 4). Далее фрагмент Ar, содержащий антигенную детерминанту (эпитоп) в комплексе с молекулой MHC II класса (I Б), транспортируется к плазматической мембране Аг-представляющей клетки, встраивается в неё и предъявляется CD4* Т-лимфоцитам.
б. Эндогенные иммуногены — продукты собственных клеток организма. Чаще всего это вирусные белки, синтезируемые вирус-инфицированными клетками хозяина, и аномальные белки опухолевых клеток. Их антигенные детерминанты предъявляются CD8+ Т-лимфоцитам в комплексе с молекулой MHC I класса (I Б).
- Антитело (AT) — гликопротеин, относящийся к классу Ig. AT специфически взаимодействует с комплементарным Ar. AT существуют в миллионах разновидностей, и каждая молекула имеет уникальный участок связывания антигенной детерминанты. AT синтезируют плазматические клетки в ходе гуморального иммунного ответа. Ig образуют один из основных классов белков крови, составляя 20% массы белка плазмы. Гены, кодирующие синтез известных классов Ig, расположены в хромосомах 2, 14 и 22.
а. Структура (рис. 11-1). Молекула Ig состоит из двух лёгких цепей (L-цепи) и двух тяжёлых цепей (Н-цепи). В цепях различают вариабельную область (V-область) в N-концевой части и постоянную, или константную область (С-область). V-область у разных AT варьирует. V-области L- и Н-цепей образуют Аг-связывающий центр, или Fab-фрагмент. Константная область молекулы Ig имеет Fc-фрагмент.
- Аг-связывающий центр образован вариабельными областями L- и Н-цепей. С Аг-связывающим центром взаимодействует антигенная детерминанта (эпитоп) иммуногена.
- Fc-фрагмент определяет специфичность связывания молекулы Ig с клетками- эффекторами (например, макрофаги, полиморфноядерные лейкоциты, тучные клетки), несущими на своей поверхности рецепторы Fc-фрагмента (рис. 11-2 и 11-4).
б. Классы Ig. В зависимости от структуры Н-цепей, выделено пять разных классов (изотипов) AT — IgA, IgD, IgE1 IgG и IgM.
- IgG — преобладающий класс AT, производится в больших количествах при иммунном (вторичном) ответе и защищает ткани от бактерий, вирусов и токсинов. IgG усиливают фагоцитоз посредством опсонизации. Из всех Ig только IgG способны проходить через плацентарный барьер.
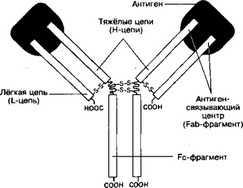
Рис. 11-1. Структура иммуноглобулина. Молекула состоит из двух идентичных тяжёлых (H) и двух идентичных лёгких (L) цепей. N-концевые области L- и Н-цепей образуют два Ar-связывающих центра. Fc-фрагмент молекулы взаимодействует со своим рецептором в мембране различных типов клеток (макрофаг, нейтрофил, тучная клетка)
(а) Фагоцитоз (рис. 11-2). IgG связываются с рецепторами Fc-фрагмента в мембране фагоцитирующих клеток, в результате чего фагоциты эффективнее поглощают и лизируют микроорганизмы.
(б) Плацентарный барьер (см. также главу 3 IX Г). На поверхности клеток трофобласта расположены рецепторы, связывающие Fc-фрагменты молекул материнских IgG. При этом связанные с рецепторами AT сначала поглощаются путём опосредованного рецепторами эндоцитоза (глава 2 I В 3 в). Далее Ig транспортируются в клетке в составе окаймлённых пузырьков (глава 2 III А 7) и выводятся из клеток трофобласта, проходят через базальную мембрану трофобласта в соединительную ткань плода и попадают в капилляры плода. Переход Ig через плаценту обеспечивает передачу пассивного иммунитета от матери к плоду.
- IgM — пентамер, пять субъединиц соединены между собой дисульфидными связями. Единственная J-цепь, связанная дисульфидными мостиками с тяжёлыми цепями, инициирует сборку пентамера. IgM — первый класс AT, продуцируемых развивающимися В-клетками при первичном попадании Ar в организм. Большая молекула IgM лггко активирует комплемент и служит как опсонин при фагоцитозе. Многие AT против грамотрицательных бактерий относятся к IgM.
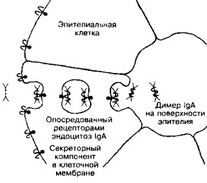
- IgA — основной класс AT в секретах (слюна, слёзы, молоко). Выделяется на поверхность слизистых оболочек, где и взаимодействует с Ar (рис. 11-3). Следовательно, IgA участвует в защитной функции организма, укрепляя барьер в слизистой оболочке пищеварительного тракта, дыхательных, половых и мочевыделительных путей. Молекула IgA в составе секрета — димер, содержащий одну J-цепь и дополнительную полипептидную цепь, называемую секреторным компонентом. Этот компонент синтезирует эпителиальная клетка, на поверхность кото-
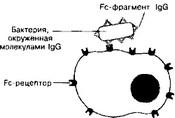
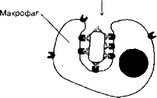 Рис. 11-2. Участие IgG в фагоцитозе.
Рис. 11-2. Участие IgG в фагоцитозе.
 Бактерия, покрытая молекулами IgG, эффективно фагоцитируется макрофагом или нейтрофилом. Fab-фрагменты IgG связываются с антигенными детерминантами на поверхности бактерии, после чего те же молекулы IgG своими Fc-фрагментами взаимодействуют с рецепторами Fc-фрагментов, расположенными в плазматической мембране фагоцита, и активируют фагоцитоз [из Alberts В et al, 1983]
Бактерия, покрытая молекулами IgG, эффективно фагоцитируется макрофагом или нейтрофилом. Fab-фрагменты IgG связываются с антигенными детерминантами на поверхности бактерии, после чего те же молекулы IgG своими Fc-фрагментами взаимодействуют с рецепторами Fc-фрагментов, расположенными в плазматической мембране фагоцита, и активируют фагоцитоз [из Alberts В et al, 1983]
Рис. 11-3. Транспорт и секреция IgA.
Молекулы IgA переносятся через эпителиальную клетку во внешнюю среду. Fc-фрагмент IgA взаимодействует со своим рецептором в мембране базальной части клетки. Образовавшийся комплекс проникает в клетку путём опосредованного рецепторами эндоцитоза. IgA отщепляется от рецептора и секре- тируется через апикальную часть эпителиальной клетки [из Alberts В et al, 1983]
рой и выделяется димер. Вероятно, секреторный компонент участвует не только в связывании молекул IgA и их внутриклеточном транспорте, но и в защите молекулы IgA от переваривания протеолитическими ферментами секретов.
- IgE специфически взаимодействует с тучными клетками и базофильными лейкоцитами (рис. 11-4). Эти клетки содержат сосредоточенные в гранулах биологически активные амины. Выделение этих веществ из клетки (дегрануляция) вызывает резкое расширение просвета венул и увеличение проницаемости их стенок. Подобную картину можно наблюдать при аллергических реакциях (например, бронхиальной астме,
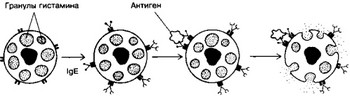
Рис. 11-4. Дегрануляция тучных клеток. На поверхности тучных клеток молекулы IgE связаны Fc-рецепторами. Ar взаимодействует с Fab-фрагментами IgE. Активированная этим сигналом тучная клетка подвергается дегрануляции [из Alberts В et al, 1983)
аллергическом рините, крапивнице). Аг-связывающие Fab-фрэгменты молекулы Ig специфически взаимодействуют с Ar, попавшим в организм. Сформированный комплекс Ar-AT взаимодействует с рецепторами Fc-фрагментов IgE, встроенных в клеточную мембрану базофила или тучной клетки. Это взаимодействие и является сигналом для экзоцитоза гистамина. IgE защищает организм от паразитов. Синтез IgE увеличивается при паразитарных инвазиях, IgE-мо- ноклональной миеломе.
- IgD. Его биологическая роль не установлена, присутствует в сыворотке в крайне низких концентрациях, появляется на поверхности развивающихся В-лимфоцитов. Б. Главный комплекс гистосовместимости. Синтез главных Ar гистосовместимости (молекул МНС) контролирует комплекс генов МНС. Гены MHC расположены в коротком плече хромосомы 6, характеризуются выраженным полиморфизмом и имеют большое количество аллелей. Спектр молекул MHC уникален для каждого организма и определяет его биологическую индивидуальность.
- Классификация. HLA-молекулы, кодируемые генами МНС, подразделяют на два класса: молекулы MHC I класса (HLA-A, HLA-B и HLA-C) и молекулы MHC II класса (HLA-D, HLA-DP, HLA-DQ и HLA-DR).
- Экспрессия. Молекулы MHC I и II классов — гликопротеины плазматической мембраны — экспрессированы на всех Аг-представляющих клетках и являются мишенями иммунного ответа при отторжении трансплантата.
а. Ar I класса представлены на поверхности практически всех клеток.
б. Ar II класса экспрессированы преимущественно на мембране иммунокомпетентных клеток, включая макрофаги, моноциты, T- и В-лимфоциты.
Аномальная экспрессия MHC II. Неиммунокомпетентные эпителиальные клетки могут аномально экспрессировать белки MHCII. Белок MHCII, экспрессируемый ¦ на поверхности эпителиальных клеток, запускает каскад патологических реакций. Примеры:
(а) фолликулярные клетки щитовидной железы — раннее проявление аутоиммунного заболевания щитовидной железы,
(б) эпителий жёлчных протоков при первичном жёлчном циррозе,
(в) р-клетки островков Лангерханса при сахарном диабете,
(г) эпителий слизистой оболочки кишечника при аутоиммунной затяжной диарее у детей.
- Строение (рис. 11-5). Каждый Ar I и II классов содержит нековалентно связанные полипептидные цепи аир. Цепи на 90% состоят из белка и на 10% из углеводов.
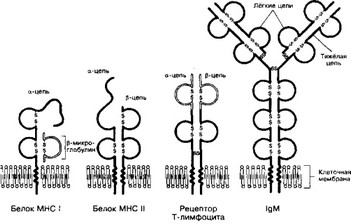
Рис. 11-5. Структура белков семейства иммуноглобулинов. Молекула MHC I класса состоит из a-цепи, внемембранная её часть связана с короткой цепью р2-микроглобулина. Молекула MHC II класса состоит из двух CE: более длинной a-цепи и p-цепи. Часть каждой цепи выступает на поверхности клеточной мембраны, цепь содержит трансмембранный участок и небольшой фрагмент в цитоплазме. Молекула рецептора Т-клеток состоит из двух цепей: аир. Каждая цепь представлена двумя внеклеточными Ig-подобными доменами, стабилизированными при помощи S-S связей, — вариабельным внеклеточным NH-концом и цитоплазматическим стабильным СООН-концом. SH-rpynna, присутствующая в цитоплазматическом фрагменте a-цепи, может взаимодействовать с мембранными или цитоплазматическими белками. Мономер молекулы IgM встраивается в плазматическую мембрану В-лимфоцитов, это рецептор Ar. Молекулярная структура рецепторов Т-клеток весьма сходна со структурой молекул MHC и Ig. Разнообразие структуры рецепторов Т-лимфоцитов и Ig обеспечивается возможностью сайт-специфической рекомбинации множества различных генных сегментов, кодирующих отдельные фрагменты молекулы [из Маррак Ф, Kannnep Д, 1986]
а. Ar I класса
- а-Цепь (44 кД) состоит из трёх доменов, формирующих спектр антигенных специфичностей молекул MHC I класса; закреплена в клеточной мембране.
- Р-Цепь (12 кД) — Р,-микроглобулин (низкомолекулярный белок, мигрирующий при электрофорезе с фракцией р-глобулинов), не кодируется МНС, не обладает полиморфизмом и не имеет трансмембранного участка.
б. Ar 11 класса
- а-Цепь (34 кД) и Р-цепь (29 кД) состоят из двух внешних доменов (аг O2 и P1, P2 соответственно), трансмембранного участка и небольшого фрагмента цепи, лежащего в цитоплазме клетки.
- Обе цепи кодируются близкорасположенными кластерами генов HLA-D региона.
- Большинство эпитопов (антигенных детерминант) HLA расположено на Р-цепи.
4. Функция. Молекулы MHC I и II класса контролируют иммунный ответ.
а. Представление Ar. Молекулы MHC II класса участвуют в представлении Ar макрофагами Т-клеткам и во взаимодействии T- и В-лимфоцитов.
б. Клеточно-опосредованный цитолиз. Молекулы MHC I и II классов распознаются поверхностноклеточными дифференцировочными Ar CD и участвуют в реакциях клеточной цитотоксичности, осуществляемой цитотоксическими Т-лимфоцитами (Tc).
- CD8, Молекулы MHC I класса взаимодействуют с молекулой CD8, экспрессируемой на мембране предшественника Tc.
- CD4. Молекулы MHC II класса взаимодействуют с молекулой CD4, экспрессируемой на мембране Т-хелпера (Тн), что вызывает выделение лимфокинов, стимулирующих пролиферацию и созревание предшественников Tc.
В. Иммунокомпетентные клетки. К ним относят T- и В-лимфоциты, NK-клетки, Аг-пред- ставляющие клетки.
- Т-лимфоциты [глава 6.1 II Б 6 е]. Только Т-клетки узнают Ar, предварительно процес- сированный и представленный на поверхности Аг-представляющих клеток. Т-лимфоци- ты (тимус-зависимые) ответственны за клеточный иммунный ответ, а также помогают реагировать на Ar В-лимфоцитам при гуморальном иммунном ответе. Т-клетки состоят из функциональных под-
Клетка-мишень
 типов CD4+ и CD8+.
типов CD4+ и CD8+.
а. Т-хелперы (Th) — CD4+ Т-клет- ки. При активации синтезируют и секретируют цитокины (ИЛ-2, ИЛ-4, ИЛ-5, ИЛ-6, у-ИФН). В ходе иммунного ответа узнают молекулы MHC II класса.
-
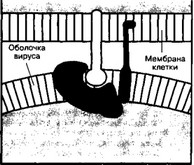 Вирус иммунодефицита человека избирательно инфицирует Т-хелперы, приводя к прогрессирующему разрушению иммунной системы и, в конечном итоге, к СПИД. С молекулой CD4 в плазматической мембране Т-хелпера связывается один из белков вирусной оболочки gpl20 (рис. 11-6). Далее мембрана Т-лимфоцита и оболочка вирусной частицы сливаются,
Вирус иммунодефицита человека избирательно инфицирует Т-хелперы, приводя к прогрессирующему разрушению иммунной системы и, в конечном итоге, к СПИД. С молекулой CD4 в плазматической мембране Т-хелпера связывается один из белков вирусной оболочки gpl20 (рис. 11-6). Далее мембрана Т-лимфоцита и оболочка вирусной частицы сливаются,
Рис. 11-6. Взаимодействие вируса иммунодефицита человека с клеткой.
Белок вирусной оболочки gpl20 связывается с молекулой CD4 мембраны лимфоцита [из Маттьюз ТД Болоньези ДП, 1988]
и генетический материал вируса оказывается внутри клетки. Если репликация вируса интенсивна, то клетка лизируется. Вирусные частицы, их фрагменты и вирусная РНК оказываются во внеклеточном пространстве.
- Образование синцития. Один из механизмов гибели клеток при ВИЧ-инфекции — образование синцитиев. Они появляются после того, как одна клетка поражается вирусом и начинает производить вирусные белки, в т.ч. белок gpl20, молекулы которого располагаются на поверхности клетки. Поскольку gpl20 обладает высоким сродством к CD4, нормальные Т-хелперы связываются и сливаются с поражённым лимфоцитом. Синцитий не функционирует и погибает. При этом уничтожается заражённая клетка и вместе с ней — множество нормальных Т-хелперов.
б. Цитотоксические Т-лимфоциты (Tc) — CD8+ Т-клетки, уничтожают инфицированные вирусом и чужеродные клетки при помощи перфорина [глава 6.1 II Б 6 е (5) (ii), глава И I Г 2]. Взаимодействуют с молекулой MHC I класса в плазматической мембране клетки-мишени.
в. Т-супрессоры (Ts) — представители CD8+ Т-клеток, регулируют интенсивность иммунного ответа, подавляя активность Th клеток. Предотвращают развитие аутоиммунных реакций. Защищают организм от нежелательных последствий иммунной реакции, от чрезмерного воспаления и аутоагрессии. Т-супрессоры обеспечивают толерантность (невосприимчивость) матери к отцовским Ar, представленным на клетках плода, что даёт возможность выживать чужеродному в иммунологическом отношении плоду в организме матери.
- В-лимфоциты (глава 6.1 II Б 6 д] ответственны за гуморальный иммунный ответ.
В мембране В-лимфоцита присутствует мономер IgM. Из красного костного мозга В-лим- фоциты мигрируют в тимус-независимые зоны лимфоидных органов. Продолжительность жизни большинства В-лимфоцитов не превышает десяти дней, если они не активируются Ar. Зрелые В-лимфоциты (плазматические клетки) вырабатывают AT — Ig всех известных классов.
- NK-клетки [глава 6.1 II Б 6 ж) не имеют поверхностных детерминант, характерных для T- и В-лимфоцитов (МНС-нерестригированные киллеры). NK-клетки убивают ауто-, алло- и ксеногенные опухолевые клетки, некоторые инфицированные вирусом и бактериями (например, Salmonella typhi) клетки. В типичных NK-клетках экспрессируются диффе- ренцировочные Ar CD2, CD56 и CD16 (рецептор Fc-фрагмента Ig).
а. Цитолиз. В отличие от цитотоксических Т-лимфоцитов, способность NK-клеток к цитолизу не связана с необходимостью распознавания молекул MHC на поверхности мишени. NK-клетки уничтожают клетку-мишень не путём фагоцитоза, а (после установления с ней прямого контакта) при помощи перфорина.
б. Гуморальная регуляция. Активность NK-клеток регулируется цитокинами. у-ИФН и ИЛ-2 усиливают цитолитическую активность NK-клеток.
в. Участие в антитело-зависимом клеточно-опосредованном цитолизе. NK-клетки, наряду с макрофагами, нейтрофилами и эозинофилами, участвуют в АТ-зависи- мом клеточно-опосредованном цитолизе. Для этого NK-клетки экспрессируют на своей поверхности рецептор Fc-фрагмента IgG (CD16). Реакция зависит от присутствия AT (Ig), узнающих клетку-мишень и связывающихся с ней. Fc-фрагмент связанных с клеткой-мишенью AT взаимодействует с рецептором Fc-фрагмента, встроенным в плазматическую мембрану NK-клетки. Природа агента, убивающего клетку-мишень в этом случае, неизвестна.
- Аг-представляющие клетки: макрофаги, В-лимфоциты, фолликулярные отростчатые клетки лимфоузлов и селезёнки, клетки Лангерханса, М-клетки в лимфатических фолликулах пищеварительного тракта, дендритные эпителиальные клетки вилочковой железы.
Эти клетки захватывают, процессируют и представляют Ar (эпитоп) на своей поверхности другим иммунокомпетентным клеткам, вырабатывают ИЛ-1 и другие цитокины, секретируют простагландин E2 (PGE2), угнетающий иммунный ответ. у-ИФН усиливает фагоцитарную и цитолитическую активность макрофагов.
Г. Взаимодействие клеток при иммунном ответе
- Гуморальный иммунный ответ. В гуморальном иммунном ответе участвуют макрофаги (Аг-представляющие клетки), Т-хелперы и В-лимфоциты.
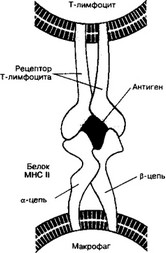
а. Макрофаг поглощает вторгшийся в организм Ar и подвергает его процессингу — расщеплению на фрагменты. Фрагменты Ar выставляются на поверхности клетки вместе с молекулой МНС. Комплекс «Аг-молекула MHC Il класса» предъявляется Т-хел- перу (рис. 11-7).
б. Т-хелпер. Узнавание Т-хелпером комплекса «Аг-молекула MHC П класса* на поверхности макрофага стимулирует секрецию ИЛ-1 (рис. 11-8). Активированный ИЛ-1 Т-хелпер синтезирует ИЛ-2 и рецепторы ИЛ-2, через которые агонист стимулирует пролиферацию Т-хелперов и цитотоксических Т-лимфоцитов. В случае Т-хелпера речь идет об аутокрин- ной стимуляции, когда клетка реагирует на тот агент, который сама же синтезирует и секретирует. Таким образом, после взаимодействия с Аг-представляющей клеткой Т-хелпер приобретает способность отвечать на действие ИЛ-2 всплеском пролиферации. Биологический смысл этого процесса состоит в накоплении такого количества Т-хелперов, которое обеспечит образование в лимфоидных органах необходимое количество плазматических клеток, способных вырабатывать AT против данного Ar.
 в. В-лимфоцит (рис. 11-8). Активация B- лимфоцита предполагает прямое взаимодействие Ar с Ig на поверхности B- клетки. В этом случае сам В-лимфоцит процессирует Ar и представляет его фрагмент в связи с молекулой MHC II на своей поверхности. Этот комплекс распознаёт Т-хелпер, отобранный при помощи того же Ar, который участвовал в отборе данного В-лимфоцита. Узнавание рецептором Т-хелпера комплекса «Аг-молекула MHC II класса*
в. В-лимфоцит (рис. 11-8). Активация B- лимфоцита предполагает прямое взаимодействие Ar с Ig на поверхности B- клетки. В этом случае сам В-лимфоцит процессирует Ar и представляет его фрагмент в связи с молекулой MHC II на своей поверхности. Этот комплекс распознаёт Т-хелпер, отобранный при помощи того же Ar, который участвовал в отборе данного В-лимфоцита. Узнавание рецептором Т-хелпера комплекса «Аг-молекула MHC II класса*
Рис. 11-7. Распознавание антигена рецептором Т-лимфоцита. Ar представлен на поверхности макрофага в связи с молекулой MHC II класса. Рецептор Т-лимфоцита распознаёт Ar только в комплексе с молекулой MHC [из Аткинсон MA, Макларен НК, 1990]
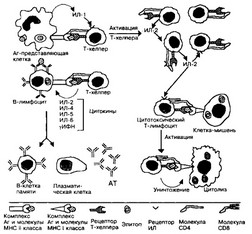
Рис. 11-8. Взаимодействие клеток при иммунном ответе. Рецептор Т-хелпера распознаёт антигенную детерминанту (эпитоп) вместе с молекулой MHC II класса, выставленные на поверхности Аг-пред- ставляющей клетки. В молекулярном взаимодействии участвует дифференцировочный Ar Т-хелпера CD4. В результате подобного взаимодействия Аг-представляющая клетка секретирует ИЛ-1, стимулирующий в Т-хелпере синтез и секрецию ИЛ-2, а также синтез и встраивание в плазматическую мембрану того же Т-хелпера рецепторов ИЛ-2. ИЛ-2 стимулирует пролиферацию Т-хелперов и активирует цитотоксические Т-лимфоциты. Отбор В-лимфоцитов производится при взаимодействии Ar с Fab-фрагментами IgM ка поверхности этих клеток. Эпитоп этого Ar в комплексе с молекулой MHC Il класса узнаёт рецептор Т-хелпера, после чего из Т-лимфоцита секретируются цитокины, стимулирующие пролиферацию В-лимфоцитов и их дифференцировку в плазматические клетки, синтезирующие AT против данного Ar. Рецептор цитотоксических Т-лимфоцитов связывается с антигенной детерминантой в комплексе с молекулой MHCI класса на поверхности вирус-инфицированной или опухолевой клетки. В молекулярном взаимодействии участвует дифференцировочный Ar цитотоксического Т-лимфоцита CD8. После связывания молекул взаимодействующих клеток цитотоксический Т-лимфоцит убивает клетку-мишень [из Gartner LP et al, 1993]
на поверхности В-лимфоцита приводит к секреции из Т-хелпера ИЛ-2, ИЛ-4, ИЛ-5 и у-ИФН, под действием которых В-клетка размножается, образуя клон плазматических клеток. В активированном В-лимфоците увеличивается количество рибосом, гранулярная эндоплазматическая сеть и комплекс Гольджи становятся более выраженными. Плазматическая клетка (рис. 11-9) синтезирует Ig. ИЛ-6, выделяемый активированными Т-хелперами, стимулирует секрецию Ig. Часть зрелых В-лимфоцитов после Аг-зависимой дифференцировки циркулирует в организме как клетки памяти.
- Клеточный иммунный ответ характеризуется пролиферацией коммитированных иммунокомпетентных клеток, реагирующих с Ar в комплексе с молекулой MHC I класса на поверхности чужеродных клеток или эндогенными иммуногенами в комплексе с молекулой MHC I класса на поверхности собственных вирус-инфицированных и опухолевых клеток. В клеточном иммунном ответе участвует цитотоксический Т-лимфоцит. Цитотоксический Т-лимфоцит (Tc). Предъявленный на поверхности клетки-мишени Ar в комплексе с молекулой MHCI класса связывается с рецептором цитотоксического Т-лимфоци- та (рис. 11-8). В этом процессе участвует молекула CD8 клеточной мембраны Tc. Секретируе- мый Т-хелперами ИЛ-2 стимулирует пролиферацию цитотоксических Т-лимфоцитов. Уничтожение клетки-мишени. Цитотоксический Т-лимфоцит распознаёт клетку- мишень и прикрепляется к ней (рис. 11-10 и 11-11). В цитоплазме активированного
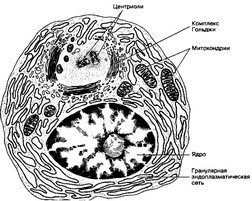
Рис. 11-9. Плазматическая клетка. Хорошо развитые гранулярная эндоплазматическая сеть и комплекс Гольджи свидетельствуют об активном синтезе и секреции белка [из Junqueira LC, Cameim J, 1991]
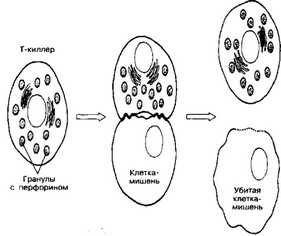
Рис. 11-10. Уничтожение клетки-мишени цитотоксическим Т-лимфоцитом. При сближении Т-лимфоцита с клеткой-мишенью после специфического взаимодействия мембранных молекул кле- ток-партнёров Т-лимфоцит убивает клетку-мишень [из Юн ДД, Кон ЖА, 1988]
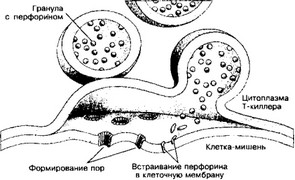
Рис. 11-11. Механизм воздействия Т-киллера на клетку-мишень. В киллере гранулы с перфорином в ответ на увеличение концентрации Ca2’сливаются с клеточной мембраной. Освободившийся перфорин встраивается в мембрану клетки-мишени с последующим образованием пор, проницаемых для воды и ионов. В результате клетка-мишень лизируется [из Юн ДД, Кон ЖА, 1988]
цитотоксического Т-лимфоцита присутствуют мелкие тёмные органеллы, напоминающие запасающие гранулы секреторных клеток. Гранулы концентрируются в той части Т-киллера, которая расположена ближе к месту контакта с клеткой- мишенью. Параллельно происходят переориентация цитоскелета и смещение в эту область комплекса Гольджи, в котором и формируются гранулы. В них содержится цитолитический белок перфорин. Выделяемые Т-киллером молекулы перфо- рина полимеризуются в мембране клетки-мишени в присутствии Ca2+. Сформированные в плазматической мембране клетки-мишени перфориновые поры пропускают воду и соли, но не молекулы белка. Если полимеризация перфорина произойдет во внеклеточном пространстве или в крови, где в избытке имеется кальций, то полимер не сможет проникнуть в мембрану и убить клетку. Специфическое действие Т-киллера проявляется только как результат тесного контакта между ним и клеткой-мишенью, который достигается за счёт взаимодействия Ar на поверхности жертвы с рецепторами Т-киллера. Сам Т-киллер защищён от цитотоксического действия перфорина. Механизм самозащиты неизвестен.
Альтернативный механизм уничтожения клетки-мишени. Существует и нашло подтверждение другое представление о механизме цитотоксического действия, согласно которому цитотоксические Т-лимфоциты и NK-клетки являются источником сигнала, который запускает уже предсуществующую суицидальную программу в клетке- мишени. Действие этого сигнала усиливают глюкокортикоиды.
Таблица 11-1. Некоторые гормоны иммунной системы, их источники и мишени
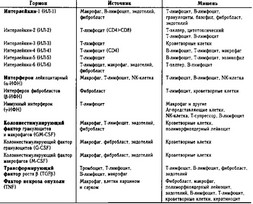
Д. Гормоны иммунной системы, их источники и мишени приведены в таблице 11-1. Цитокины. Ранее продуцируемые лимфоцитами гормоны именовали лимфокинами, а моноцитами — моноки- ны. В настоящее время для большой группы гуморальных факторов, секретируемых клетками системы тканей внутренней среды, принят термин «цитокины».
Е. Реакции гиперчувствительности. Иммунные механизмы, обеспечивающие защиту организма, могут приводить к повреждению органов и тканей, реализуясь в виде реакций гиперчувствительности. Классификация Джелла и Кумбса подразделяет гиперчувствительность на четыре основных типа (в зависимости от механизмов, участвующих в их реализации). Многие иммунопатологические процессы опосредованы комбинацией нескольких реакций гиперчувствительности.
- Реакции гиперчувствительности I типа (немедленного типа, атопические, реагино- вые). При данном типе реакций происходит взаимодействие Ar с IgE1 приводящее к высвобождению биологически активных медиаторов (главным образом, гистамина) из тучных клеток и базофилов. Примеры реакций I типа — поллиноз, бронхиальная астма, анафилактический шок.
- Реакции гиперчувствительности II типа. AT (обычно IgG или IgM) связываются с Ar на поверхности клеток, что активирует фагоцитоз и приводит к развитию АТ-зависи- мого клеточно-опосредованного цитолиза. Реакции сопровождаются активацией комплемента СЗ с последующим фагоцитозом клеток или активацией всей системы комплемента с последующим цитолизом и повреждением ткани. Пример реакции II типа — аутоиммунная гемолитическая анемия.
- Реакции гиперчувствительности III типа (иммунных комплексов). Комплексы, образованные Ar и соответствующим AT, активируют систему комплемента, приводя к развитию воспалительной реакции. Пример реакции III типа — острый гломерулонефрит.
- Реакции гиперчувствительности IV типа (клеточно-опосредованные или замедленного типа). В этих реакциях принимают участие не AT, а Т-клетки, взаимодействующие с соответствующим Ar (сенсибилизированные Т-клетки). Сенсибилизированные Т-клетки после связывания соответствующего Ar оказывают либо непосредственное цитотоксическое действие на клетки-мишени, либо их цитотоксический эффект опосредуется с помощью лимфокинов. Примеры реакций IV типа — контактный дерматит и реакция отторжения трансплантата.
Источник: Под ред. Э.Г. Улумбекова, Ю.А. Челышева, «Гистология (введение в патологию)» 1997