
В конце 60-х — начале 70-х годов XX в. были разработаны лабораторные методы клонирования клеток in vitro. Для этого понадобилось создать питательные среды, материалы для лабораторной посуды и термостаты-инкубаторы, позволившие имитировать условия внутренней среды организма для сохранения жизнеспособности клеток млекопитающих. В течение 10—12 лет удавалось клонировать только опухолевые клетки, поскольку их собственным свойством является способность неограниченно (в благоприятных для них внешних условиях) делиться митозом. Г. Келлер и Ц. Милыптейн в 1974—1975 гг. применили метод получения гибридных соматических клеток, который использовали цитогенетики для изучения локализации генов, контролирующих тот или иной признак в той или иной хромосоме (гибридные клетки выбрасывают большую часть хромосом, но не все), к лимфоцитам, но с иными целями. Г. Келлер и Ц. Милыптейн получили гибридные клетки из лимфоидной опухоли (миелома) и нормального лимфоцита. Гибридные клетки имели часть хромосом (а следовательно, и свойств) нормального лимфоцита (другая часть хромосом выбрасывалась из клеток в течение первых делений, пока геном не стабилизировался) и часть — от опухоли. Размножались только клетки, унаследовавшие от миеломы способность к неограниченному делению. Параллельно в них шел биосинтез тех или иных продуктов нормального лимфоцита, например антител; последние и требовались Г. Келлеру и Ц. Милыптейну. Неограниченно делящиеся клетки «позволяют» себя клонировать, т.е. физически «рассадить» по одной (каждую в отдельную посуду) и получить клоны клеток — потомков одной клетки. Такие клетки назвали гибридомами.
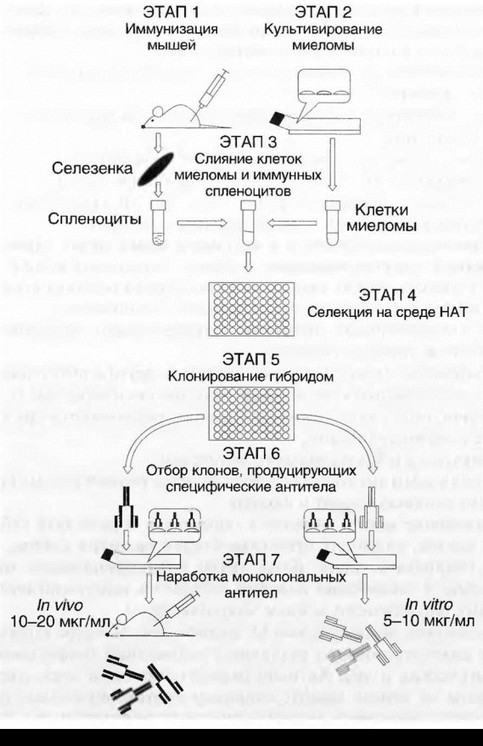
Лимфоциты для гибридизации получают из селезенки или лимфоузлов, предварительно иммунизированных целевым антигеном мышей (чаще всего — линии Balb/c). В качестве опухолевых клеток- партнеров используют специально выведенные для получения гибридом мутантные клетки мышиной миеломы (иногда ее же называют плазмоцитомой, что в данном случае одно и то же), полученные из перевивной линии миеломных клеток МОРС-21 от мышей Balb/c,
поддерживаемой в культуре in vitro с 1921 г. Келлер и Милыптейн использовали мутантные миеломные клетки, выведенные цитогенетиками ранее также для получения гибридных клеток млекопитающих, но в иных целях: поскольку гибридные клетки выкидывают большую часть хромосом, по оставшимся цитогенетики устанавливали локализацию тех или иных признаккодирующих генов в конкретных хромосомах.
«Мутантность» миеломных клеток необходима для метаболической селекции клеток гибридом от неслившихся с лимфоцитами клеток миеломы. Эта «мутантность» состоит в отсутствии в миеломных клетках действующего гена, кодирующего фермент ГГФРТ. Данный фермент катализирует синтез гуанина (одного из 4 азотистых оснований, входящих в состав ДНК) из гипоксантина. Вскоре после первой публикации Келлера и Милыптейна в распоряжении «гибридомщи- ков» во всем мире оказались две удобные линии мутантных миелом — P3/X63-Ag8-653 и SP2/0-Agl4, которые быстро пролиферируют и сами не продуцируют никаких частей иммуноглобулинов.
Суспензию лимфоидных клеток от иммунных мышей смешивают в одной пробирке в минимальном объеме среды с суспензией миеломных клеток и на 1—2 мин добавляют сливающий агент. У Келлера и Милыптейна сначала этим агентом был, как и у цитогенетиков, вирус Сендай, но через несколько месяцев уже все «гибридомщики» в качестве сливающего агента использовали синтетические полимеры — полиэтиленгликоли с молекулярными массами от 1540 до 6000 D. По прошествии 1—2 мин суспензию клеток, содержащую смесь неслившихся лимфоидных клеток, неслившихся клеток миеломы и гибридных клеток 3 вариантов («лимфоцит—лимфоцит», «миелома—миелома», «лимфоцит—миелома»; из них искомыми клетками являются только гибриды «лимфоцит—миелома»), отмывают и разводят в рассчитанном объеме селективной среды HAT. HAT означает «Hypoxanthine-Aminopterin Thymidine». Указанные 3 компонента в известных концентрациях вводят в полную культуральную среду. В течение первых 7—10 дней культивирования названной смеси клеток в культуре происходит следующее:
избирательно блокирующего ферменты биосинтеза пиримидиновых оснований de novo из М5М10-метилен-тетрагидрофолата; биосинтез пуриновых оснований из гипоксантина в этих клетках также невозможен в связи с отсутствием ГГФРТ;
Основные этапы гибридомной технологии показаны на рис. 5.1.
От клеток миеломы данные гибридные клетки наследуют свойство неограниченной пролиферации. От нормальных иммунных В-лимфоцитов — биосинтез иммуноглобулинов.
Пролиферация «non-stop» позволяет клонировать гибридные клетки, т.е. рассеять по 1 в лунку и подождать, когда из этой одной клетки при благоприятных условиях культивирования вырастет клон, т.е. много одинаковых клеток (с точностью до спонтанных мутаций). Какие из получившихся «лимфоцит—миеломных» гибридных клеток продуцируют заданные антитела, выясняют, отбирая из лунок пробы супернатанта на соответствующий иммуноанализ. В дальнейшем избранные гибридомы неоднократно реклонируют и выводят в массовые культуры — реакторы in vitro или асцитные опухоли у сингенных мышей. Из культуральных супернатантов или асцитных жидкостей выделяют гибридомные моноклональные антитела в очищенном виде.
Если клон клеток гибридомы синтезирует антитела, то эти антитела называют моноклональными. Как показал опыт, все клетки клонированной гибридомы синтезируют одинаковые антитела — и по специфичности активного центра, и по изотипу тяжелой цепи, т.е. моноклональные антитела — не только продукт моноклона, но и препарат одинаковых иммуноглобулинов. В конце 1970-х годов научились выращивать in vitro и клонировать Т-лимфоциты, не скрещенные с опухолевой линией клеток. Это стало возможным только после открытия фактора роста Т-лимфоцитов, позже названного ИЛ-2. Именно клонирование Т-лимфоцитов позволило открыть субпопуляции CD4+ Т-лимфоцитов и сделать множество других открытий, в том числе идентифицировать ВИЧ (достаточные для исследования количества генов и белков вируса смогли наработать
только на Т-лимфоцитах, культивируемых in vitro в присутствии факторов роста). В-лимфоциты без превращения их в гибридомы можно заставить делиться (что позволяет их клонировать), если инфицировать их вирусом Эпштейна—Барр (он трансформирует нормальные В-лимфоциты в опухоль из В-лимфоцитов).
Таким образом, моноклональные антитела, продуцируемые одним клоном, являются:
Стабильные культуры гибридом способны продуцировать в неограниченном количестве моноклональные антитела.
Моноклональные антитела в настоящее время широко применяют в различных областях медицины. В основе применения монАТ в различных областях лежит возможность получения больших количеств высокоаффинных антител, специфичных по отношению:
Применение монАТ позволяет определять и разделять субпопуляции клеток, различать отдельные стадии развития клеток, более точно типировать ткани, более точно идентифицировать микроорганизмы, а также более надежно определять иммунологическими методами биологически важные макромолекулы.
В медицине на основе монАТ разработано большое количество систем диагностирования различных заболеваний (инфекционных, онкологических и др.). Активно разрабатываются и лекарственные препараты на основе монАТ, например противоопухолевые препараты, антицитокиновые антитела (препарат ремикейт и др.) Однако было показано, что применение мышиных монАТ для иммунотерапии опухолевых заболеваний приводит к появлению различных побочных эффектов и отличается слабой эффективностью действия.
Модификация мышиных монАТ (гуманизированные антитела и различные конъюгаты монАТ — с радионуклидами, лекарствами, токсинами и пр.) открывает новые возможности в диагностике и лечении опухолевых и других заболеваний.
Лимфоциты для гибридизации получают из селезенки или лимфоузлов, предварительно иммунизированных целевым антигеном мышей (чаще всего — линии Balb/c). В качестве опухолевых клеток- партнеров используют специально выведенные для получения гибридом мутантные клетки мышиной миеломы (иногда ее же называют плазмоцитомой, что в данном случае одно и то же), полученные из перевивной линии миеломных клеток МОРС-21 от мышей Balb/c,
поддерживаемой в культуре in vitro с 1921 г. Келлер и Милыптейн использовали мутантные миеломные клетки, выведенные цитогенетиками ранее также для получения гибридных клеток млекопитающих, но в иных целях: поскольку гибридные клетки выкидывают большую часть хромосом, по оставшимся цитогенетики устанавливали локализацию тех или иных признаккодирующих генов в конкретных хромосомах.
«Мутантность» миеломных клеток необходима для метаболической селекции клеток гибридом от неслившихся с лимфоцитами клеток миеломы. Эта «мутантность» состоит в отсутствии в миеломных клетках действующего гена, кодирующего фермент ГГФРТ. Данный фермент катализирует синтез гуанина (одного из 4 азотистых оснований, входящих в состав ДНК) из гипоксантина. Вскоре после первой публикации Келлера и Милыптейна в распоряжении «гибридомщи- ков» во всем мире оказались две удобные линии мутантных миелом — P3/X63-Ag8-653 и SP2/0-Agl4, которые быстро пролиферируют и сами не продуцируют никаких частей иммуноглобулинов.
Суспензию лимфоидных клеток от иммунных мышей смешивают в одной пробирке в минимальном объеме среды с суспензией миеломных клеток и на 1—2 мин добавляют сливающий агент. У Келлера и Милыптейна сначала этим агентом был, как и у цитогенетиков, вирус Сендай, но через несколько месяцев уже все «гибридомщики» в качестве сливающего агента использовали синтетические полимеры — полиэтиленгликоли с молекулярными массами от 1540 до 6000 D. По прошествии 1—2 мин суспензию клеток, содержащую смесь неслившихся лимфоидных клеток, неслившихся клеток миеломы и гибридных клеток 3 вариантов («лимфоцит—лимфоцит», «миелома—миелома», «лимфоцит—миелома»; из них искомыми клетками являются только гибриды «лимфоцит—миелома»), отмывают и разводят в рассчитанном объеме селективной среды HAT. HAT означает «Hypoxanthine-Aminopterin Thymidine». Указанные 3 компонента в известных концентрациях вводят в полную культуральную среду. В течение первых 7—10 дней культивирования названной смеси клеток в культуре происходит следующее:
- неслившиеся лимфоциты и гибриды «лимфоцит—лимфоцит» погибают в силу своей природной недолговечности;
- неслившиеся клетки миеломы и гибриды «миелома—миелома» погибают от невозможности осуществлять биосинтез своей ДНК в присутствии аминоптерина — метаболического яда,
избирательно блокирующего ферменты биосинтеза пиримидиновых оснований de novo из М5М10-метилен-тетрагидрофолата; биосинтез пуриновых оснований из гипоксантина в этих клетках также невозможен в связи с отсутствием ГГФРТ;
- единственные клетки, имеющие возможность выжить в среде HAT, это искомые гибридные клетки «лимфоцит—миелома»: биосинтез пуриновых оснований у них обеспечен ГГФРТ, ген которой получен из нормального лимфоцита, и средовым гипоксантином, а биосинтез пиримидиновых оснований осуществляется из средового тимидина с участием тимидинкиназы.
Основные этапы гибридомной технологии показаны на рис. 5.1.
От клеток миеломы данные гибридные клетки наследуют свойство неограниченной пролиферации. От нормальных иммунных В-лимфоцитов — биосинтез иммуноглобулинов.
Пролиферация «non-stop» позволяет клонировать гибридные клетки, т.е. рассеять по 1 в лунку и подождать, когда из этой одной клетки при благоприятных условиях культивирования вырастет клон, т.е. много одинаковых клеток (с точностью до спонтанных мутаций). Какие из получившихся «лимфоцит—миеломных» гибридных клеток продуцируют заданные антитела, выясняют, отбирая из лунок пробы супернатанта на соответствующий иммуноанализ. В дальнейшем избранные гибридомы неоднократно реклонируют и выводят в массовые культуры — реакторы in vitro или асцитные опухоли у сингенных мышей. Из культуральных супернатантов или асцитных жидкостей выделяют гибридомные моноклональные антитела в очищенном виде.
Если клон клеток гибридомы синтезирует антитела, то эти антитела называют моноклональными. Как показал опыт, все клетки клонированной гибридомы синтезируют одинаковые антитела — и по специфичности активного центра, и по изотипу тяжелой цепи, т.е. моноклональные антитела — не только продукт моноклона, но и препарат одинаковых иммуноглобулинов. В конце 1970-х годов научились выращивать in vitro и клонировать Т-лимфоциты, не скрещенные с опухолевой линией клеток. Это стало возможным только после открытия фактора роста Т-лимфоцитов, позже названного ИЛ-2. Именно клонирование Т-лимфоцитов позволило открыть субпопуляции CD4+ Т-лимфоцитов и сделать множество других открытий, в том числе идентифицировать ВИЧ (достаточные для исследования количества генов и белков вируса смогли наработать
только на Т-лимфоцитах, культивируемых in vitro в присутствии факторов роста). В-лимфоциты без превращения их в гибридомы можно заставить делиться (что позволяет их клонировать), если инфицировать их вирусом Эпштейна—Барр (он трансформирует нормальные В-лимфоциты в опухоль из В-лимфоцитов).
Таким образом, моноклональные антитела, продуцируемые одним клоном, являются:
- высокоспецифичными, направленными к заданной антигенной детерминанте;
- идентичными по изотипу, аллотипу и идиотипу, а также по аффинитету и физико-химическим характеристикам.
Стабильные культуры гибридом способны продуцировать в неограниченном количестве моноклональные антитела.
Моноклональные антитела в настоящее время широко применяют в различных областях медицины. В основе применения монАТ в различных областях лежит возможность получения больших количеств высокоаффинных антител, специфичных по отношению:
- к иммуногенным антигенам, определяющим гистосовместимость и дифференцировку;
- дифференцировочным, опухолевым и другим антигенам клеточной поверхности, которые лишены полиморфизма и неиммуногенны в аллогенных системах, но распознаются при ксеногенной иммунизации;
- вирусным и бактериальным антигенам;
- единичным антигенным детерминантам разнообразных белков, нуклеиновых кислот и сахаров.
Применение монАТ позволяет определять и разделять субпопуляции клеток, различать отдельные стадии развития клеток, более точно типировать ткани, более точно идентифицировать микроорганизмы, а также более надежно определять иммунологическими методами биологически важные макромолекулы.
В медицине на основе монАТ разработано большое количество систем диагностирования различных заболеваний (инфекционных, онкологических и др.). Активно разрабатываются и лекарственные препараты на основе монАТ, например противоопухолевые препараты, антицитокиновые антитела (препарат ремикейт и др.) Однако было показано, что применение мышиных монАТ для иммунотерапии опухолевых заболеваний приводит к появлению различных побочных эффектов и отличается слабой эффективностью действия.
Модификация мышиных монАТ (гуманизированные антитела и различные конъюгаты монАТ — с радионуклидами, лекарствами, токсинами и пр.) открывает новые возможности в диагностике и лечении опухолевых и других заболеваний.