
Большинство микроорганизмов, подобно высшим растениям, способны синтезировать все аминокислоты, входящие в состав клеточных белков. Исходным материалом для построения углеродных скелетов аминокислот служат промежуточные продукты обмена углеводов в реакциях гликолиза и цикла трикарбоновых кислот. Наиболее важными из них являются кетокислоты. Перевод азота в органические соединения осуществляется через аммиак. Нитраты, нитриты, молекулярный азот восстанавливаются до аммиака в результате ассимиляционной нитратредукции и включаются в состав органических соединений.
Некоторые аминокислоты образуются путем прямого аминировакия кетокислот аммонием. Аминогруппы вводятся путем прямого аминирования или транса минирования. Таким путем
происходит первичная ассимиляция аммиака и построение первичных аминокислот, которые служат источником аминных групп для образования других аминокислот путем переаминирования, т. е. переноса аминогрупп с аминокислот на кетокислоты. В реакциях прямого аминирования принимают участие глутаматдегидрогеназа и аланиндегидрогеназа. Так, а- кетоглутаровая кислота при участии глутаматдегидрогеназы реагирует с аммонием, образуя глутаминовую кислоту, аланин- дегидрогеназа катализирует включение аммония в пиро- виноградную кислоту и образование аланина; аспартатаммиаклиаза катализирует синтез аспарагиновой аминокислоты:
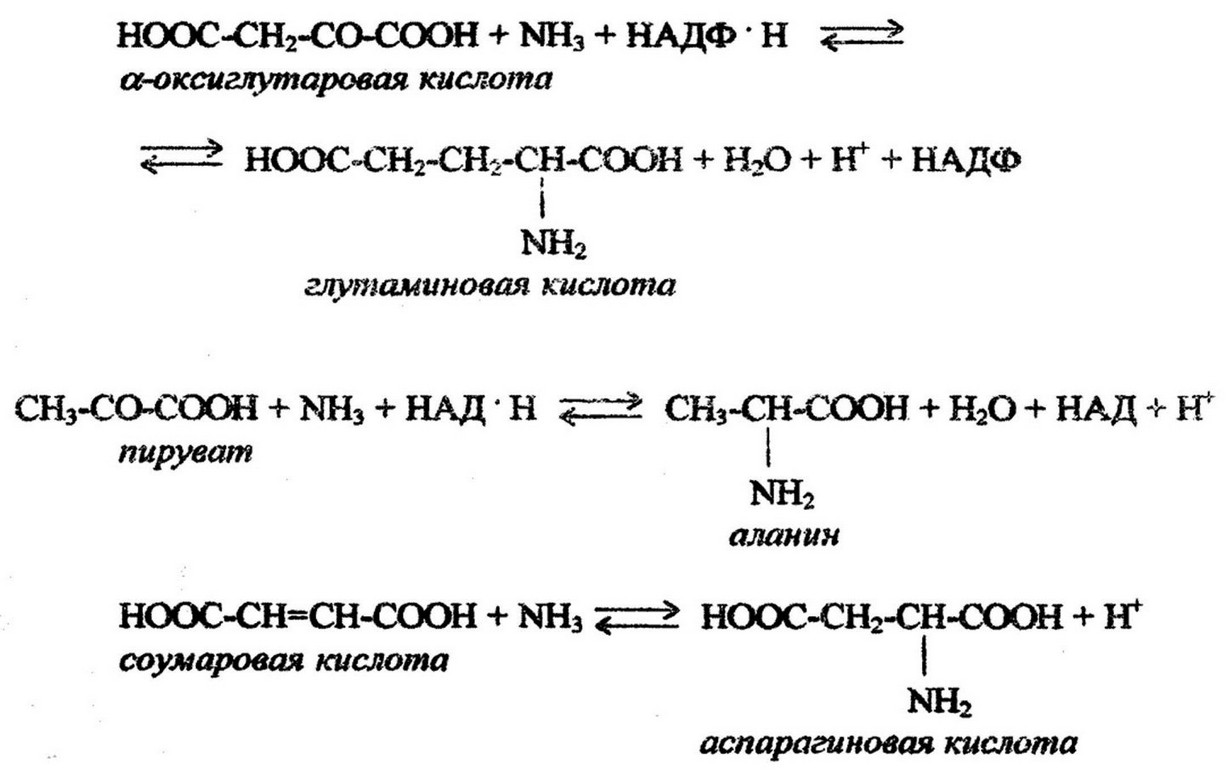
Многие гетеротрофные бактерии образуют новые аминокислоты в основном через трансаминирование или переаминирование аминокислот, потребляемых из питательной среды. Важным в биосинтетических реакциях является превращение одних аминокислот в другие: например, глутамата в аргинин и пролин; аспарагиновой кислоты в лизин, метионин, треонин; треонина в глюкокол и т. д.
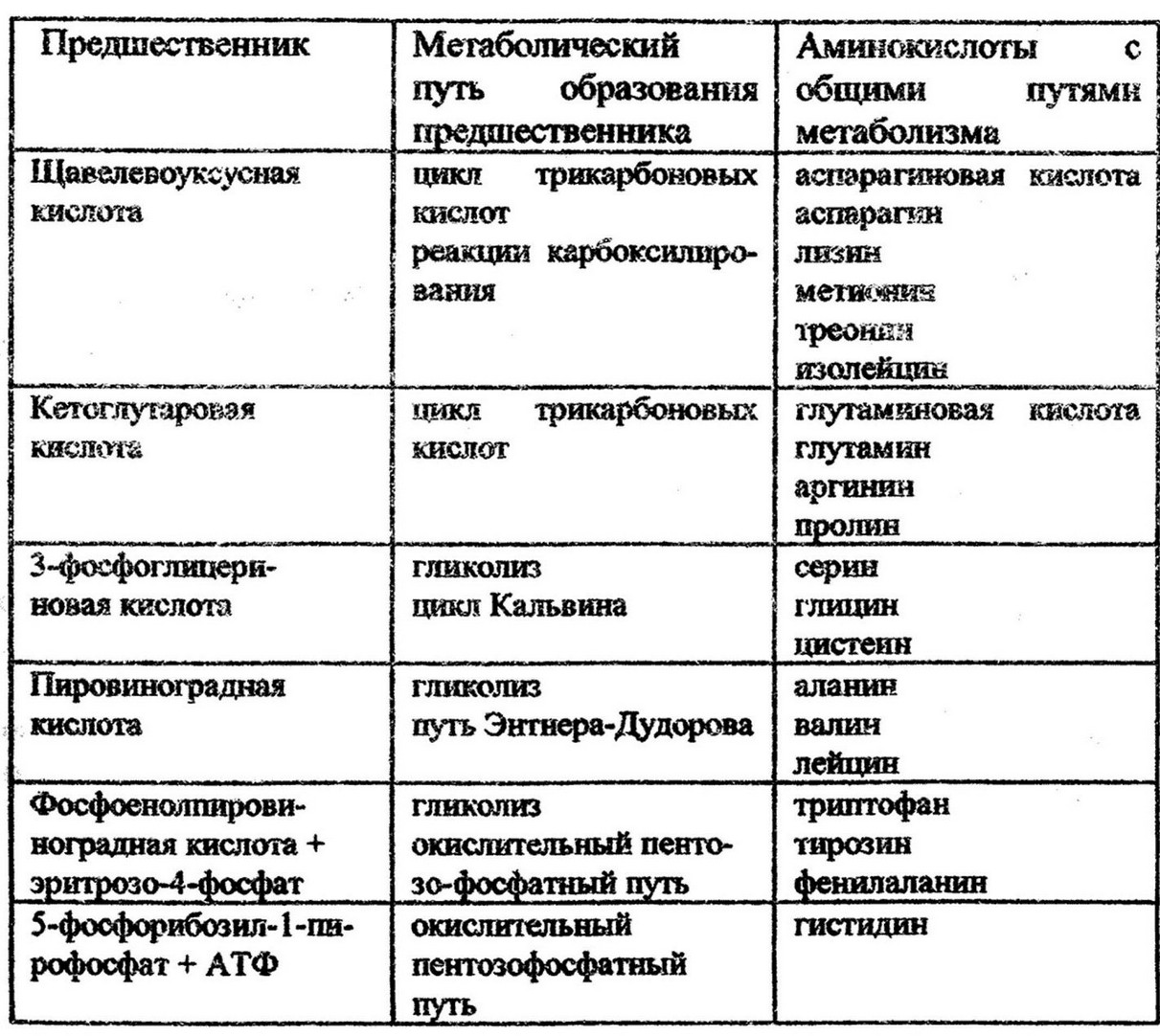
Особенностью биосинтеза аминокислот является наличие общих метаболических путей. Всего выделяют шесть таких путей и соответственно им все 20 аминокислот основного набора разделяют на шесть биосинтетических семейств (табл.З). Из них только гистин имеет самостоятельный путь биосинтеза.
Пути биосинтеза аминокислот приводят также к образованию иных азотсодержащих соединений, таких как полиамины, пурины, фолиевая кислота и другие вещества, необходимые для клетки.
Таблица 3
Особенности биосинтеза аминокислот
- Биосинтез белков
Аминокислоты, синтезированные внугриклеточно или потребленные из внешней среды, полимеризуются в молекулу белка. Механизм полимеризации довольно сложный и осуществляется в несколько этапов при непосредственном участии всех видов нуклеиновых кислот, которые играют решающую роль в биосинтезе белка. В ДНК закодирована информация о структуре каждого типа белка, характерного для определенного организма. Различные участки ДНК функционально неоднородны, и одна ее молекула может определять синтез большого числа функционально и химически различных белков клетки. Участок ДНК (несколько пар нуклеотидов), определяющий синтез одного типа белка, обозначается как ген. Ген передает информацию особой РНК3 которая синтезируется на нем как на матрице и называется информационной, или матричной, РНК (мРНК). При синтезе мРНК копируется нуклеотидная последовательность информации, закодированной ь ДНК, называемая транскрипцией. Процесс перевода нуклеотидной последовательности мРНК в последовательность аминокислот в полипептиде называется трансляцией. Таким образом, синтез полипептида включает два процесса - транскрипцию и трансляцию:
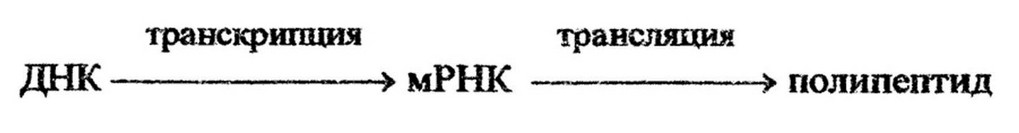
Включение конкретной аминокислоты в синтезируемую пептидную цепь определяется комбинацией трехсоседних нуклеотидов в молекуле мРНК - триплетом.
Триплеты ДНК называются кодонами. Большинство аминокислот кодируется несколькими триплетами. Это значит, что генетический код вырожден (табл.41
Генетический код
Таблица 4
|
Первая |
Вторая буква 1 |
|||
|
буква |
У |
ц |
А |
г ! |
|
У |
УУУФен1 |
УЦУСер |
УАУТир |
УГУ Цис |
|
|
УУЦФен |
УЦЦСер |
УАЦТир |
УГЦЦис |
|
|
УУА Лей |
УЦА Сер |
УАА (нет)2 |
УГА (нет)2 |
|
|
УУГ Лей |
УЦГСер |
УАГ (нет)2 |
УПГТрн |
|
Ц |
ЦУУ Лей |
ПДУ Про |
ЦАУ Гис |
ЦГ У Apr |
|
|
ЦУЦ Лей |
ЦЦЦПро |
ЦАЦ Гис |
ЦГЦАрг |
|
|
ЦУА Дей |
ЦЦАПро |
ЦААГли |
ЦТ A Apr |
|
|
ЦУГ Лей |
ЦЦГПро |
ЦАГГли |
ЦГТ Apr |
|
А |
АУУ Иле |
АЦУТре |
ААУ Асв |
АГУ Сер |
|
|
АУЦ Иле |
АЦЦТре |
ААЦ Асн |
АЩСер |
|
|
АУА Иле |
АДА Тре |
АААЛиз |
АГА Apr |
|
|
ДУГ Мег |
АЦГТре |
ААГ Лиз |
ААГ Apr |
|
.. Г |
ГУ У Вал |
ЩУ Ала |
ГАУ Асп |
ГГУГлн 1 |
|
|
ГУЦ Вал |
ГЦЦАда |
ГАД Асп |
ГТЦ Гт \ |
|
1 |
ГУАЬал |
ГИД Ала |
ГААГлу |
ГГ А Г ли |
|
|
ГУГ Вал |
ГЦГ Ала |
ГАГ Гду |
ПТ Гли 1 |

Все молекулы мРНК содержат сигналы начала и конца кодируемого ими белка. Матричная РНК связывается с малой субчастицей рибосомы, объединяя группу рибосом в полисомы.
Синтез белка у всех организмов происходит на рибосомах (рис.7.8). Доставку аминокислот к рибосомам осуществляют транспортные РНК (тРНК) Каждая аминокислота доставляется к рибосомам специфичной для нее тРНК.
Первым этапом синтеза белка является активирование

Рис. 7.8. Схема биосинтеза белка
аминокислот и образование аминоацил - тРНК. Этот процесс катализируется специфичными для каждой аминокислоты ферментами аминоацил - тРНК-сннтетазами при участии АТФ в соответствии со следующим уравнением:
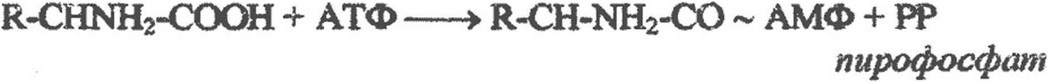
Транспортные РНК переносят аминокислоты к рибосомам, где они пептидными связями соединяются в пептидную цепь.
Перенос аминокислот к рибосомам посредством тРНК необходим, потому что сами по себе аминокислоты не способны узнавать кодоны в мРНК. Их узнают тРНК благодаря наличию в составе своей молекулы антикодонов. Кроме того, присоединение аминокислоты к рибозному остатку тРНК активирует карбоксильную группу аминокислоты и она способна к образованию пептидной связи. Узнавание кодона антикодоном (тРНК) контролируется рибосомами. Рибосомы движутся вдоль мРНК в направлениисчитывая кодоны путем
присоединения к ним соответствующей аминоацил-тРНК. С каждым присоединением аминокислоты рибосома передвигается на один триплет вдоль мРНК, «считывая» шаг за шагом всю информацию.
Когда рибосома доходит до последнего триплета, то после присоединения соответствующей ему аминокислоты к растущей полипеитидной цепи синтез белка прекращается.
Процесс трансляции, а следовательно, и образования полипегггидной цепи, очень сложен, В нем различают три стадии: инициацию, элонгацию и терминацию.
Инициация синтеза всех белков начинается со связывания N -фор мил "Метионил - тРНК с пептидильным центром (П) 30S-cy6- частицы рибосомы, которая присоединена к инициирующему триплету мРКК (АУГ или ГУГ). Затем нужная аминоamp;цил-тРНК подходит к аминоацильному центру (А) 50S-субчастицы рибосомы и образуется первая пептидная связь, Таким образом, стадия нжщиации включает все реакции, обеспечивающие формирование пептидной связи между первыми двумя аминокислотами.
Далее следует стадия элонгации. Ока включает реакции связывания соответствующих аминоацил-тРНК, образование пептидных связей и транслокацию. Пептидная цепь растет в направлении от Н-концевой аминокислоты к С-концевой, т. е. от аминокислоты со свободней аминогруппы к аминокислоте со свободной карбоксильной группой.
На стадии терминации полностью синтезированный полипептид освобождаете® от концевой тРНК, Рибосомы отделяются от мРНК к диссоциируют на исходные субчастицы gt; Терминацию синтеза белка осуществляют так называемые факторы освобождена. Это белки (RFi-'l, RF-2), способные узнавать терминирующие кодоны (УАГ, УДА, УГА). Связывание фактора осгзобождения с тер ми и ир ующн м кодоном активирует
пептидилтрансферазу и она гидролизует связь между полипептидом и тРНК.
На рибосомах синтезируется белок первичной структуры, под которой понимают последовательность аминокислот в пептидной цепи. После завершения синтеза полипептидная цепь свертывается, приобретая вторичную и третичную структуру, а а ряде случаев объединяется с* другими полипептидными цепями и в результате образуется белок четвертичной структуры. Синтез белка происходит с расходованием большого количества энергии. Только для
образования одной пептидной связи затрачивается количество энергии, эквивалентное 4 молекулам АТФ.
- Биосинтез липидов
Микроорганизмы способны осуществлять биосинтез как сложных, так и простых липидов, которые входят в состав клеточной стенки и цитоплазматической мембраны, являются запасными веществами, компонентами пигментных систем и цепей электронного транспорта.
Источником для синтеза липидов микроорганизмами служат углеводы, спирты, органические кислоты. Кроме них, необходимо наличие в среде фосфатов для образования фосфор ил ированных предшественников биосинтеза.
Образованию липидов предшествует синтез эфиров жирных кислот и коэнзима А. Затем следуют реакции конденсации жирных кислот с глицерином, приводящие к появлению липидов. Пути синтеза липидов сложные, реакции катализируются многими ферментами и протекают с затратой значительных количеств энергии. Например, на синтез только стеариновой кислоты из глюкозы затрачивается 945,7 кал.
Глюкоза может служить также источником глицерина:
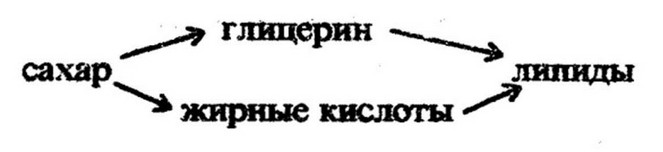
Глицерин получается в процессе анаэробного расщепления углеводов путем восстановления глицеринового альдегида. Жирные кислоты образуются из промежуточных продуктов распада углеводов, главным образом уксусной кислоты, уксусного альдегида, пировиноградной кислоты и этилового спирта. Важнейшей из них является уксусная кислота:
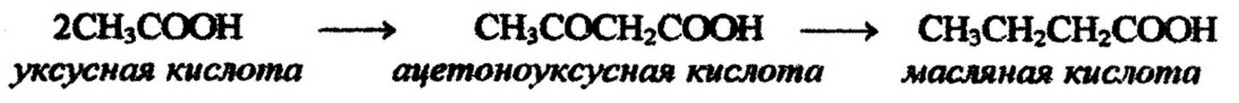
Но не сама уксусная кислота участвует в биосинтезе липидов, а ее производное - ацетилкоэнзим А, который является источником ацетильных радикалов и занимает центральное положение в метаболизме липидов:
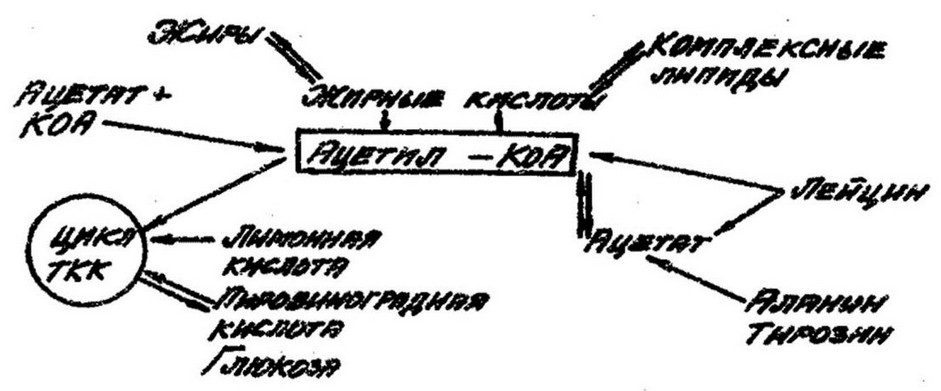
Макроэргическая тиоэфирная связь а цетил-К оА при
гидролизе высвобождает S200 кал. Данная энергия используется для переноса ацетильных остатков, необходимых для биосинтеза жирных кислот. Ацетилкоэнзим А за счет различных органических кислот при помощи ряда циклических процессов - конденсирования, гидрогенизации и дегидратации - обеспечивает образование длинных цепей жирных кислот, являющиеся компонентами лшшдов в клетках эубактерий.
У цианобактерий выявлены пали ненасыщенные жирные кислоты, имеющие две и более двойных связей, для образования которых аэробным бактериям необходим молекулярный кислород.
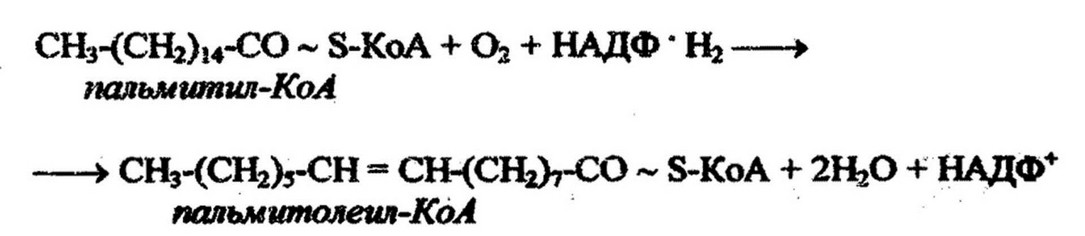
Второй путь введения двойной связи в молекулу кислоты при ее синтезе осуществляется у облигатно анаэробных и некоторых аэробных эубактерий в результате реакции дегидратации.
Синтез фосфолипидов состоит из нескольких этапов. Исходный субстрат фосфодиоксиацетон (промежуточный продукт гликолитического пути) восстанавливается до 3-фосфоглицерина к
которому присоединяются два остатка жирных кислот с образованием фосфатидной кислоты. Происходит активирование ее с помощью ЦТФ и присоединение к фосфатной группе серина, инозита, глицерина приводят к синтезу фосфатидил серина, фосфатидилинозита и фосфатидилглицерина.