
Периодической называют культуру , в которую после засева бактериями соответствующей питательной среды в процессе роста не добавляются никакие компоненты и не удаляются из нее продукты метаболизма (закрытая система). Закономерность
размножения бактериальной популяции в периодической культуре описывается s-образной кривой, которая выражает зависимость числа клеток (или биомассы) от продолжительности роста культуры.
При развитии популяции в жидкой среде различают четыре основные фазы роста, последовательно сменяющие друг друга: начальную, или лаг-фазу; экспоненциальную, или логарифмическую фазу; стационарную фазу и фазу отмирания (рис. 4.1).
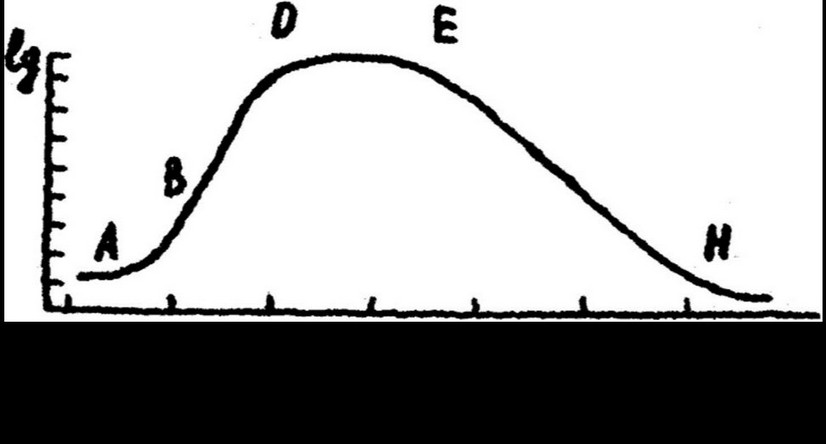
Лаг-фаза охватывает период времени от засева бактерий в питательную среду до начала деления клеток. В течение лаг-фазы число клеток не увеличивается. Происходит приспособление клеток к новым условиям среды, выражающееся в синтезе индуцибельных ферментов, повышение метаболической активности, увеличение размеров клеток.
Длительность лаг-фазы зависит от качества среды, в которой ранее выращивалась культура и в которую произведен посев. Чем полноценнее среда, в которую засевается культура, тем короче лаг- фаза. Большое внимание на продолжительность лаг-фазы оказывает количество и возраст засеваемых клеток. Чем старше культура и чем меньше взято клеток для посева, тем длительнее эта фаза. Лаг-фаза может затянуться, если питательная среда не полноценная, т. е. если
i
она не содержит в достаточном количестве аминокислот, витаминов и других необходимых веществ.
Иногда в культуре может наблюдаться наличие двух лаг-фаз. Это обычно происходит на средах, содержащих смесь питательных веществ. Некоторые культуры на таких средах проявляют способность к поочередному использованию источников питания. В 1942 г. Моно впервые описал поочередное использование двух типов углеводов Е. coli и назвал это явление диауксией. В основе этого явления лежит катаболитная репрессия. При развитии Е. coli на среде, содержащей глюкозу и сорбит, вначале потребляется глюкоза; синтез ферментов, необходимых для потребления сорбита, угнетается. Они образуются только после того, как вся глюкоза будет потреблена и клетка приступит к использованию другого источника питания - сорбита. В случае диауксии лаг-фаза снова повторяется.
Потребление субстрата сопровождается увеличением бактериальной массы - биомассы. Прирост биомассы называется урожаем. Урожай клеток отражает интенсивность развития культуры. Важным показателем, характеризующим эффективность использования субстрата или продуктивность культуры в конкретных условиях среды, является экономический коэффициент. Он выражается отношением урожая (выросшей биомассы) к
количеству потребленного субстрата:
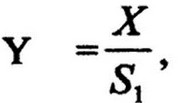 где Y -
где Y -
экономический коэффициент, X - количество выросшей биомассы, г; Si - количество потребленного субстрата, г. Количество потребленного субстрата определяют как разность между исходной S0 и остаточной S концентрацией его:
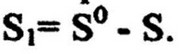
Лаг-фаза сменяется экспоненциальной, или логарифмической фазой роста. В этой фазе все клетки находятся в состоянии активного деления. Нарастание клеток идет в геометрической прогрессии: скорость размножения постоянная; продолжительность генерации, т. е. время между двумя последовательными делениями клетки, минимальная. Прирост бактерий в единицу времени пропорционален наличному количеству клеток.
Показателем интенсивности роста культуры является удельная скорость роста (р), которая означает прирост биомассы, или числа
клеток за один час на единицу растущей биомассы. Математически это выражается формулой:
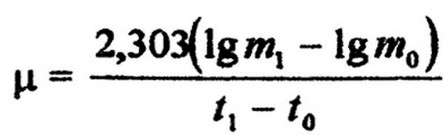
где Шо и mi - величины начальной и через время t биомассы (или числа клеток);
 - промежуток времени между начальным и
- промежуток времени между начальным и
последующим определениями биомассы или подсчетом числа клеток.
Другим важным параметром скорости роста культуры является среднее время удвоения биомассы, или время генерации (g), т. е. время между двумя последовательными делениями клетки. Оно определяется по увеличению численности клеток за определенный промежуток времени t. Если в начале экспоненциальной фазы число клеток было А, то после первой генерации, т. е. после первого деления количество клеток удвоится и будет равно А * 2; после второй генерации количество клеток станет
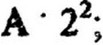 после третьей -
после третьей -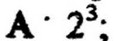 . после п генераций конечное число клеток В
. после п генераций конечное число клеток В
окажется.
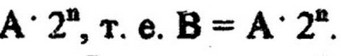
В микробиологической практике для выражения общего числа клеток чаще пользуются не абсолютными числами, так как они достигают огромных величин, а их логарифмами. Прологарифмировав данное уравнение, получим:
IgB = IgA + и lg2
Величины А и В (начальное и конечное число клеток) определяем одним из методов подсчета числа клеток: в счетной камере, на мембранных фильтрах или путем высева культуры из чашки Петри со средой с последующим учетом выросших колоний.
Число генераций п находим из уравнения:
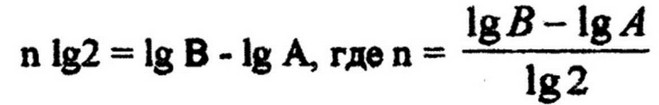
Зная число генераций (делений клеток) и продолжительность опыта, легко определить скорость размножения клеток (в минутах):
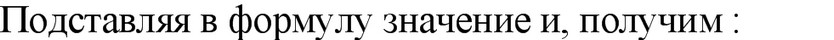
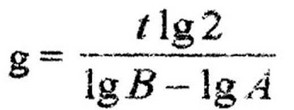
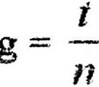
Учитывая, что даже в экспоненциальной фазе происходит отмирание клеток (около 20 %), в данную формулу вносится 20 %~ ная поправка, что от 2 составляет 0,4. В конечном итоге формула получит следующий вид:
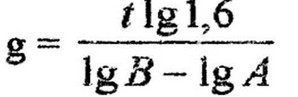
Удельная скорость роста и время генерации взаимосвязаны Поэтому, определив экспериментально один из параметров, можно вычислить другой по формуле:
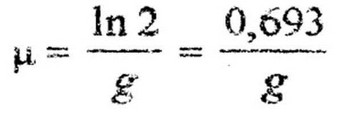
Продолжительность скорости размножения ш
экспоненциальной фазе различна у разных микроорганизмов Так, для Е. coli она может быть 20-30 мин; для азотобактера - 70-80 мин; для клубеньковых бактерий - до 90 мин. Рост культуры в экспоненциальной фазе называют сбалансированным, считал, что в это время все компоненты клетки синтезируются в одинаковой степени. Однако б условиях периодической культуры рост является сбалансированным в течение очень короткого интервала времени В процессе роста культуры происходит изменение состава
питательной среды; питательные вещества потребляются, а ненужные продукты метаболизма накапливаются. Реакцией клетки на изменения качества среды является изменение интенсивности синтеза РНК, белка, полисахаридов и других необходимых компонентов.
Качественные изменения среды являются основной причиной замедления роста культуры. В результате наступает стационарная фаза. Она соответствует периоду, когда число жизнеспособных клеток в популяции перестает увеличиваться, хотя многие из них находятся в стадии активного деления. В этой фазе количестве вновь появляющихся и переходящих в стадию покоя клеток
приблизительно равно. На протяжении всей фазы численность популяции не изменяется. У разных видов она наступает через различные промежутки времени: у Е. coli через 24 ч, у азотобактера через 72 ч. Количество клеток в стационарной фазе достигает максимума, размеры их становятся близкими к размерам клеток исходного посевного материала. После достижения стационарной фазы жизнеспособность клеток снижается и начинается фаза отмирания.
Таким образом, цикл развития периодической культуры довольно сложный. Действующие на клетку факторы очень многочисленны и меняются по ходу развития культуры. Вначале культура страдает от высокой концентрации питательных веществ по сравнению с их содержанием в среде, с которой взят посевной материал, т. е. происходит торможение роста избытком субстрата. Через некоторое время культура начинает развиваться с возрастающей скоростью, достигая ее максимального уровня. Затем постепенно сказывается исчерпание элементов питания и отравление продуктами обмена, развитее культуры замедляется и конце концов прекращается.
-ч