
Среди отделов коры большого мозга, которые первыми стали известны физиологам, были зрительные области. Дженнари (Jennari) в 1776 г. описал зрительную кору. Теперь ее размеры и строение на макро- и микроуровне хорошо изучены. Выяснены также некоторые физиологические свойства зрительной коры, но декодирующий процесс как целое, т. е. сложная функция зрительного восприятия, полностью еще не раскрыт. Однако те немногие детали этого сложного механизма, которые стали известны в последние годы (главным образом благодаря обширным микрофизиологическим исследованиям), обнадеживают и побуждают к дальнейшему изучению.
Зрительная кора. При изучении корковой проекционной зоны сетчатки применялись почти все методы исследования, упомянутые в гл. 1. У обезьян эта зона обнаружена в заднем отделе
затылочной доли; анатомические, нейрохирургические и неврологические данные говорят о том, что в головном мозгу человека она занимает то же положение (рис. 14). Центральная ямка представлена участком в несколько квадратных сантиметров на наружной, выпуклой поверхности затылочной доли. При стимуляции одной точки сетчатки светом или толчками электрического тока вызванные потенциалы возникают в нескольких, по меньшей мере в двух, точках коры.
В отличие от сравнительно однородной организации латерального коленчатого тела зрительная кора состоит из несколь-

коры (справа) и на срезах латерального коленчатого тела таламуса (слева). Участки коры, отмеченные крестиками и мелкими точками, соответствуют белым участкам в коленчатом теле.
ких слоев. Волокна от коленчатого тела оканчиваются в четвертом (от поверхности коры) из шести различимых под микроскопом слоев. Обширная сеть волокон передает отсюда информацию к соседним слоям. От третьего и пятого слоев отходит множество волокон, идущих к подкорковым нейронам и соседним областям коры.
Примечательная особенность этой системы состоит в том, что вертикальных, колончатых связей между отдельными слоями гораздо больше, чем горизонтальных, боковых связей. Из такой организации следует, что корковыми проекциями отдельных рецептивных полей служат ограниченные вертикальные колонки. Эта колончатая организация была подтверждена элект- рофизиологическими данными.
Ориентационные колонки. Недавно Хыобел и Визель (Hubei, Wiesel) установили, что все нейроны этих кортикальных колонок отвечают на простые линейные стимулы одного типа, т. е. на прямые световые линии (щели или границы контраста), идущие в определенном направлении. Например, если кошке показать вертикальную линию на белом поле, то от соответствующих клеток коры отводятся высокочастотные потенциалы действия.



 Рис. 15. Колончатая организация клеток в области корковой проекции сетчатки (вверху) и схема ориентационно-специфического ответа «простых» корковых клеток (внизу). Клетка, специфически отвечающая потенциалами действия на вертикальные стимулы, не отвечает иа стимуляцию горизонтальной линией, Частота ответа растет по мере того, как линия поворачивается и приближается к вертикальному положению.
Рис. 15. Колончатая организация клеток в области корковой проекции сетчатки (вверху) и схема ориентационно-специфического ответа «простых» корковых клеток (внизу). Клетка, специфически отвечающая потенциалами действия на вертикальные стимулы, не отвечает иа стимуляцию горизонтальной линией, Частота ответа растет по мере того, как линия поворачивается и приближается к вертикальному положению.
Но если ориентацию линии изменить, электрический ответ этих клеток исчезнет (рис. 15). Клетки других колонок реагируют иа линии с иной ориентацией. На этом основании в качестве основной аналитической единицы постулирован набор ориентационных колонок, содержащих все специальные разновидности в пределах 180° (гиперколонка). В то же время оказалось, что сложность ответов варьирует с расстоянием от поверхности коры, что указывает на существование нескольких слоев «простых» и более «сложных» клеток.
«Простые» и «сложные» клетки. Нейроны, отвечающие на простые линейные стимулы (щели, края или темные полосы), получили название «простых», а те, которые отвечают на стимулы сложной конфигурации и на движущиеся стимулы, были названы «сложными».
On- и off-клетки тоже можно обнаружить в зрительной коре, но здесь они расположены не концентрически. Эти два вида клеток резко разграничены, и направление разделяющей их границы зависит от ориентационной специфичности нейрона (рис. 16). Предполагается, что функция ориентационных колонок осуществляется путем латерального торможения.
Распределение колончатых функциональных единиц в зрительной коре неоднотипно. Показано, что «простые» клетки преобладают в поле 17 по Бродману, которое лежит в центре зрительной коры, а в полях 18 и 19 чаще встречаются «сложные» клетки. Высказано предположение, что информация, проанализированная и переработанная простыми клетками, передается
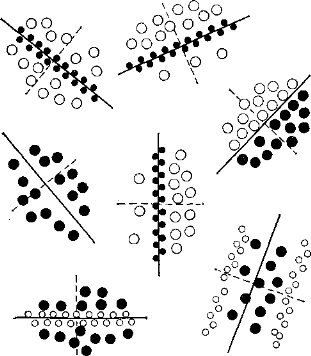
Рис. 16. Различная ориеитация рецептивных полей «простых» корковых нейронов. Оп-клетки (белые кружки) резко отграничены от off-клеток (черные кружки). Направление разделяющей их линии соответствует ориентации, к которой чувствителен данный «простой» нейрон. Диаметр рецептивного поля составляет у кошки около 1 мм.
сложным для дальнейшего анализа. Таким образом, весьма вероятно, что зрительное восприятие — результат кооперации разных клеток.
Информация, закодированная, проанализированная и переработанная в сетчатке и таламусе, декодируется нескольким» миллионами нейронов в коре. В конечном результате, таким образом, получается аналоговое сообщение в соответствии с принципом отражения внешней среды.
«Сверхсложные» нейроны. В зрительной коре описан также третий класс нейронов, а именно «сверхсложные» клетки. Это истинные интегрирующие единицы, выполняющие в зрительной системе функцию синтеза. Они получают информацию от нижележащих нейронов и обеспечивают единообразие восприятия пространства и формы.
Восприятие цвета. Хотя эти электрофизиологические данные создают прочную основу для дальнейшего изучения работы мозга, их недостаточно для интерпретации восприятия в понятиях психологии. Множество вопросов все еще остается без ответа. Как на сравнительно простой пример укажем на то, что мы почти ничего не знаем о цветовоспринимающей функции мозга. Кажется несомненным, что сообщение о цвете кодируется до- минаторно-модуляторной системой сетчатки, но нет никаких данных о перерабатывающей и декодирующей активности в латеральном коленчатом теле и коре. В таламусе обезьяны были исследованы нейроны разных типов, отвечающие на различные цветовые стимулы, но полученные результаты противоречивы. Скудность данных объясняется тем, что лабораторные млекопитающие (грызуны, кошки, обезьяны) плохо различают цвета. Собака, например, совсем их не различает. Большая часть информации получена в опытах на лягушках. И хотя эти опыты выполнены весьма остроумно и имеют большое значение, их результаты нельзя перенести на человека.
Зрительная кора. При изучении корковой проекционной зоны сетчатки применялись почти все методы исследования, упомянутые в гл. 1. У обезьян эта зона обнаружена в заднем отделе
затылочной доли; анатомические, нейрохирургические и неврологические данные говорят о том, что в головном мозгу человека она занимает то же положение (рис. 14). Центральная ямка представлена участком в несколько квадратных сантиметров на наружной, выпуклой поверхности затылочной доли. При стимуляции одной точки сетчатки светом или толчками электрического тока вызванные потенциалы возникают в нескольких, по меньшей мере в двух, точках коры.
В отличие от сравнительно однородной организации латерального коленчатого тела зрительная кора состоит из несколь-
коры (справа) и на срезах латерального коленчатого тела таламуса (слева). Участки коры, отмеченные крестиками и мелкими точками, соответствуют белым участкам в коленчатом теле.
ких слоев. Волокна от коленчатого тела оканчиваются в четвертом (от поверхности коры) из шести различимых под микроскопом слоев. Обширная сеть волокон передает отсюда информацию к соседним слоям. От третьего и пятого слоев отходит множество волокон, идущих к подкорковым нейронам и соседним областям коры.
Примечательная особенность этой системы состоит в том, что вертикальных, колончатых связей между отдельными слоями гораздо больше, чем горизонтальных, боковых связей. Из такой организации следует, что корковыми проекциями отдельных рецептивных полей служат ограниченные вертикальные колонки. Эта колончатая организация была подтверждена элект- рофизиологическими данными.
Ориентационные колонки. Недавно Хыобел и Визель (Hubei, Wiesel) установили, что все нейроны этих кортикальных колонок отвечают на простые линейные стимулы одного типа, т. е. на прямые световые линии (щели или границы контраста), идущие в определенном направлении. Например, если кошке показать вертикальную линию на белом поле, то от соответствующих клеток коры отводятся высокочастотные потенциалы действия.
Рис. 15. Колончатая организация клеток в области корковой проекции сетчатки (вверху) и схема ориентационно-специфического ответа «простых» корковых клеток (внизу). Клетка, специфически отвечающая потенциалами действия на вертикальные стимулы, не отвечает иа стимуляцию горизонтальной линией, Частота ответа растет по мере того, как линия поворачивается и приближается к вертикальному положению.
Но если ориентацию линии изменить, электрический ответ этих клеток исчезнет (рис. 15). Клетки других колонок реагируют иа линии с иной ориентацией. На этом основании в качестве основной аналитической единицы постулирован набор ориентационных колонок, содержащих все специальные разновидности в пределах 180° (гиперколонка). В то же время оказалось, что сложность ответов варьирует с расстоянием от поверхности коры, что указывает на существование нескольких слоев «простых» и более «сложных» клеток.
«Простые» и «сложные» клетки. Нейроны, отвечающие на простые линейные стимулы (щели, края или темные полосы), получили название «простых», а те, которые отвечают на стимулы сложной конфигурации и на движущиеся стимулы, были названы «сложными».
On- и off-клетки тоже можно обнаружить в зрительной коре, но здесь они расположены не концентрически. Эти два вида клеток резко разграничены, и направление разделяющей их границы зависит от ориентационной специфичности нейрона (рис. 16). Предполагается, что функция ориентационных колонок осуществляется путем латерального торможения.
Распределение колончатых функциональных единиц в зрительной коре неоднотипно. Показано, что «простые» клетки преобладают в поле 17 по Бродману, которое лежит в центре зрительной коры, а в полях 18 и 19 чаще встречаются «сложные» клетки. Высказано предположение, что информация, проанализированная и переработанная простыми клетками, передается
Рис. 16. Различная ориеитация рецептивных полей «простых» корковых нейронов. Оп-клетки (белые кружки) резко отграничены от off-клеток (черные кружки). Направление разделяющей их линии соответствует ориентации, к которой чувствителен данный «простой» нейрон. Диаметр рецептивного поля составляет у кошки около 1 мм.
сложным для дальнейшего анализа. Таким образом, весьма вероятно, что зрительное восприятие — результат кооперации разных клеток.
Информация, закодированная, проанализированная и переработанная в сетчатке и таламусе, декодируется нескольким» миллионами нейронов в коре. В конечном результате, таким образом, получается аналоговое сообщение в соответствии с принципом отражения внешней среды.
«Сверхсложные» нейроны. В зрительной коре описан также третий класс нейронов, а именно «сверхсложные» клетки. Это истинные интегрирующие единицы, выполняющие в зрительной системе функцию синтеза. Они получают информацию от нижележащих нейронов и обеспечивают единообразие восприятия пространства и формы.
Восприятие цвета. Хотя эти электрофизиологические данные создают прочную основу для дальнейшего изучения работы мозга, их недостаточно для интерпретации восприятия в понятиях психологии. Множество вопросов все еще остается без ответа. Как на сравнительно простой пример укажем на то, что мы почти ничего не знаем о цветовоспринимающей функции мозга. Кажется несомненным, что сообщение о цвете кодируется до- минаторно-модуляторной системой сетчатки, но нет никаких данных о перерабатывающей и декодирующей активности в латеральном коленчатом теле и коре. В таламусе обезьяны были исследованы нейроны разных типов, отвечающие на различные цветовые стимулы, но полученные результаты противоречивы. Скудность данных объясняется тем, что лабораторные млекопитающие (грызуны, кошки, обезьяны) плохо различают цвета. Собака, например, совсем их не различает. Большая часть информации получена в опытах на лягушках. И хотя эти опыты выполнены весьма остроумно и имеют большое значение, их результаты нельзя перенести на человека.