Генерация и проведение импульсов
В процессе филогенеза возникли специальные органы рецепции и передачи информации о внешних стимулах. Рецепторы представляют собой специализированные структуры, преобразующие внешние стимулы в электрохимические сигналы — импульсы, которые затем передаются по нервным проводящим путям в сенсорную кору. Рецепторы тесно соприкасаются с соответствующими нервными клетками (нейронами), точнее с их отростками, которые проводят импульсы, возникающие в рецепторах, к телу клетки (соме). Основная функция нейрона состоит в передаче импульсов, генерируемых рецепторами. Разумеется, эта функция варьирует в различных нейронах животного и человеческого организма: одни нейроны способны порождать ритмические импульсы, другие анализируют и интегрируют импульсы, которые к ним приходят. Здесь мы будем рассматривать только роль нейронов как передатчиков импульсов. Упрощая, можно сказать, что генерацию импульсов осуществляют рецепторные нейроны, их передачу — остальные нейроны. Рассмотрим обе эти функции.
Чтобы понять распространение коротких, периодических электрохимических импульсов, надо сначала обратиться к нейрону в состоянии покоя. Обычно разность потенциалов между внешней и внутренней сторонами мембраны рецепторного нейрона в покое составляет примерно от 30 до 100 мВ. Эту разность мы называем потенциалом покоя. Ее можно измерить, введя в клетку микроэлектрод толщиной около 1 мкм, а второй электрод поместив с внешней стороны клеточной мембраны. Разность потенциалов регистрируется достаточно чувствительным прибором. Пока еще не установлено, чем вызывается эта разность. Но одно можно сказать с уверенностью: внутриклеточная жидкость (жидкое содержимое клетки) по своему составу отличается от наружной, т. е. внеклеточной, жидкости. В то время как внутри клетки в избытке имеются ионы калия, в тканевой жидкости больше ионов натрия. Содержимое клетки богато белком, а в наружной жидкости много ионов хлора. Как полагают, высокую концентрацию калия и низкую концентрацию натрия в клетке создает работа так называемого «натриевого насоса» — особого активного метаболического процесса. Потенциал покоя, согласно Ходжкину (Hodgkin), достаточно хорошо объясняется неравномерным распределением ионов.
Аналоговый ответ. При раздражении рецепторного нейрона потенциал покоя снижается на 30—50 мВ, т. е. происходит частичная деполяризация. В таком состоянии достаточно второго стимула гораздо меньшей интенсивности, чтобы потенциал покоя исчез. Упомянутое снижение потенциала в рецепторе строго локально — оно происходит только в том участке клеточной
Потенциал поноя Рецепторный потенциал Потенциал действия
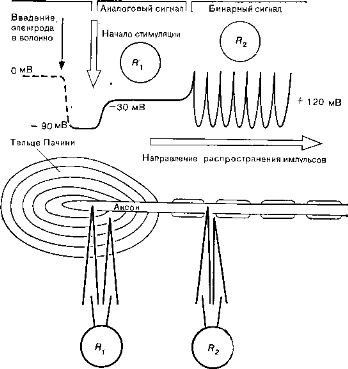
Рис. 1. Генерация и распространение нервного импульса в тельце Пачини и в аксоне, выходящем из рецептора. Rx и fe - положение регистрирующих микроэлектродов (внизу) и соответствующие записи (вверху): рецепторный потенциал и периодический потенциал действия.
мембраны, на который подействовал стимул. Еще одна особенность деполяризации состоит в том, что степень изменения потенциала зависит от интенсивности стимула: более слабые стимулы лишь ненамного снижают потенциал покоя, а сильные вызывают более выраженную деполяризацию. Можно сказать, что это локальное снижение потенциала пропорционально внешнему стимулу; поэтому в терминах кибернетики его можно назвать аналоговым ответом.
Такое локальное уменьшение разности потенциалов, носящее аналоговый характер, биофизики называют генераторным потенциалом. Генераторный потенциал рецептора некоторые авторы называют рецепторным потенциалом. Этот местный сдвиг потенциала представляет собой очень важный феномен, так как он порождает нервные импульсы в аксоне, связанном с рецепторной клеткой. Нервный импульс не остается локальным,
он распространяется и, как мы увидим, не является аналоговым, а носит — опять-таки на языке кибернетики — характер бинарного (двоичного) сигнала. Тельце Пачини. Рецепторный потенциал часто исследуют в тельцах Пачини, которые в большом числе имеются в коже и других тканях (например, в брюшине) человека и животных.
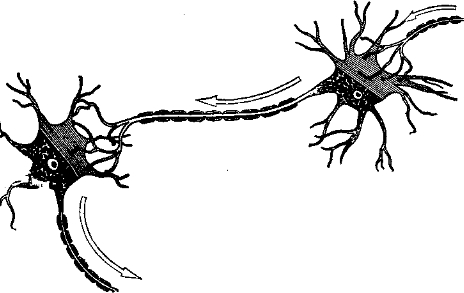
Рис. 2. Схема строения нейрона. Видно, как к верхнему нейрону подходят окончания аксона от другого нейрона. Короткие отростки—дендриты, длинные волокна с миелиновой оболочкой — аксоны (обратите внимание иа разрывы в оболочке, называемые перехватами Ранвье). Заштрихованные участки тела клетки изображают клеточную мембрану с сидящими на ней концевыми вздутиями. В остальной части клетки показаны ядро, ядрышко и гранулы тигроида. Эти последние играют важную роль в метаболизме и синтезе белка. Стрелками показано направление распространения импульсов.
Тельце Пачини по своей структуре напоминает луковицу; оно реагирует на давление, т. е. на механическую силу, изменяющую его структуру (рис. 1). В нервном окончании,погруженном в тельце Пачини, возникает рецепторный потенциал, после чего сигнал распространяется к другим частям нейрона в форме периодических импульсов.
Строение нейрона показано на рис. 2. В головном мозгу и остальной нервной ткани большинства животных тела нейронов имеют звездчатую или слегка округленную форму. Наиболее заметная особенность нейрона — наличие отростков, похожих на корешки. Короткие ветвящиеся отростки называются денд- ритамщ их функция обычно состоит в приеме сигнала. Длинный одиночный отросток, часто снабженный миелиновой оболочкой, называется аксоном. Функция его состоит в проведении нервных импульсов. В то время как некоторые нейроны лишены дендритов, ни одна нервная клетка ни у одного вида животных не может не иметь аксона. Аксой (или нервное волокно) — это самая важная для проведения импульсов структура, основной элемент нервных проводящих путей.
Бинарный ответ. Если рецепторный потенциал превысит порог возбуждения аксона, тотчас же произойдет его полная деполяризация. Это внезапное изменение называют потенциалом действия. Как мы видели, в рецепторе потенциал покоя только снижается. В аксоне же он совсем исчезает; более того, на одну тысячную секунды — или даже на меньший интервал — внутренняя часть волокна становится слегка электроположительной по отношению к окружающей жидкости.
Такая «реверсия» потенциала периодически повторяется. В момент максимума потенциала действия (спайка) мембрана аксона становится нечувствительной к последующим стимулам (рефрактерность), и новый импульс возникает лишь после того, как она восстановит свое нормальное состояние поляризации. Таким образом получается характерный для нервного проведения поток импульсов. Серия спайков распространяется вдоль нервного волокна, в миелинизированном волокне перескакивая с одного перехвата Ранвье (см. рис. 2) на другой. В отличие от местной деполяризации при возникновении импульсов деполяризация не варьирует с изменением интенсивности стимула. Будет ли пришедший сигнал пороговым или гораздо более сильным, амплитуда спайка остается одинаковой (ответ типа «всё или ничего»).
Два типа потенциала — два типа мембраны. Из сказанного выше видно, что генерация нервных импульсов в рецепторных структурах проходит две фазы. Первая фаза — это местная частичная деполяризация, создающая рецепторный потенциал. Когда это снижение потенциала достигает достаточной величины, деполяризуется также мембрана примыкающего аксона и
по нерву распространяется серия спайков (потенциалов действия). Различия в свойствах рецепторного потенциала и потенциала действия указывают на возможное различие свойств мембраны в соответствующих участках:
Возникает вопрос: каким образом внешний стимул преобразуется рецептором в периодические импульсы? Или, говоря на языке кибернетики, каким образом в рецепторе происходит преобразование аналогового сигнала в бинарный? Как мы видели, рецепторный потенциал отражает интенсивность и длительность стимула: деполяризация пропорциональна силе входного сигнала, и она длится, пока длится стимуляция. Именно поэтому электрическую реакцию рецептора можно назвать аналоговым ответом, который отражает параметры стимула.
Место преобразования. Нервные сигналы типа «всё или ничего» мы находим при изучении электрических изменений в любой части периферического афферентного нервного пути. Отсюда можно сделать вывод, что преобразование аналоговых сигналов в бинарные, т. е. кодирование, совершается в рецепторе. Затем нервные сигналы передаются центральным нейронам в этой преобразованной форме, которая облегчает передачу, а возможно, и хранение (см. гл. II).
Информация. Под информацией обычно понимают сообщения или новые сведения о каком-нибудь событии или изменении. В этом смысле нервные импульсы, идущие от периферических рецепторов в головной мозг, передают информацию точно так же, как импульсы, бегущие по телефонному кабелю, или радиоволны. Чем меньше вероятность события, тем большую ценность представляет информация о нем. С помощью теории вероятности информацию можно измерить и придать ей численное выражение. Единицей при измерении количества информации, содержащейся в сообщении, служит бит. Это количество информации получается при единичном выборе типа «да или нет», если вероятность события равна 0,5. Теория информации интересуется только количественными сторонами информации, оставляя в стороне ее смысл и значение для получателя,
В отличие от этого нейрофизиология имеет дело также с качеством информации, т. е. с той пользой, которую организм может извлечь из полученного сообщения для своего благополучия и приспособления.
Кодирование. Сообщения передаются в нервной системе, как вообще в коммуникационных системах, в форме сигналов (кодов). Преобразование в определенную систему сигналов (кодирование) облегчает передачу и хранение информации. Необходимое условие состоит в том, чтобы сообщение могло быть декодировано в конечном воспринимающем звене. В случае нервной системы можно сказать, что кодирование происходит в рецепторах, а декодирование — в головном мозгу.
Бинарный код. Кодирующие системы, в которых используются только два вида сигналов (0 и 1), называются бинарными (двоичными)! В нервной системе информация, вероятно, передается с помощью такого бинарного кода. Во всяком случае, на это указывает правило «всё или ничего» для нервных сигналов, состоящих из серии импульсов, за которыми следуют реф* рактерные периоды.
Бинарные сигналы применяются для передачи и хранения информации также в компьютерах. Использование двоичной системы счисления, предложенное Нейманом (Neumann) и другими, удобно, так как позволяет применять простую и надежную электронную аппаратуру.
Информационный канал. Для передачи информации требуется соответствующая среда. Это то, что мы называем каналами (рис. 3). Информационными каналами служат, например, телефонные кабели и аксоны нервных клеток. Пропускную способность таких каналов определяют как максимальное количество информации, которое может быть передано в единицу времени; ее выражают числом битов в секунду. Сигналы, поступающие в систему извне, называются входными, а сигналы, выходящие на другом ее конце, — выходными. На практике часть информации при передаче теряется в результате искажения входных сигналов под влиянием различных факторов (так называемого шума), мешающих передаче сообщений в нервной системе. Вопрос о такого рода искажении, в частности в перехватах Ранвье по ходу миелинизированных нервных волокон и в других местах, составляет еще не решенную нейрофизиологическую проблему. Возможно, что искажения импульсации — бинарных сигналов, передаваемых по нервному волокну,— корректируются сигналами в других, параллельных волокнах, передающими ту же самую информацию. Как указывает Нейман, использование параллельных зашумленных каналов статистически дает благоприятные результаты. Таким образом, сравнительно слабая изоляция нервных волокон в конечном итоге улучшает функцию нервного пути посредством статистической
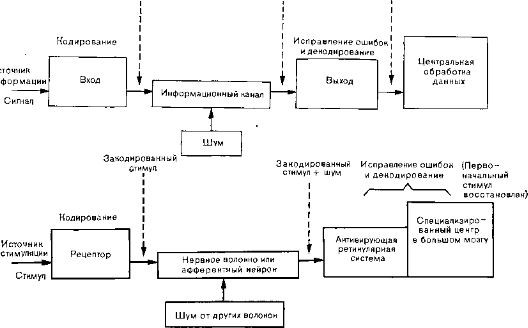
Закодированный Закодированный Первоначальный сигнал сигнал + шум сигнал
Рис. 3. Схема передачи информации в технических коммуникационных системах (вверху) и в нервной системе (внизу).
корреляции серий импульсов, идущих по нескольким каналам.
Кодирующий механизм рецепторов пока еще мало изучен. Два типа активности мембраны у аксона, связанного с рецепторной клеткой, указывают на то, что перекодирование должно происходить в месте соединения участков мембраны того и другого типа. В механорецепторах (например, в рецепторе давления — тельце Пачини) критической точкой является первый перехват Ранвье по ходу миелинизированного волокна, идущего от рецепторной клетки (см. рис. 1).
Сальтаторное проведение. Бинарный импульс, идущий по нервному волокну с миелиновой оболочкой, перескакивает от перехвата к перехвату. Такое сальтаторное проведение выявлено в эксперименте. Показано, что порог возникновения импульса в аксоне очень низок именно в перехватах Ранвье. Хаксли и Стемпфли (Huxley, Stampfli) отводили микроэлектродами потенциал действия от участка нерва между двумя перехватами и нашли, что на всем отрезке волокна между двумя перехватами потенциалы действия появляются одновременно без задержки в проведении. Такая задержка обнаруживается только между двумя соседними перехватами (рис. 4).
Изоляция нервных волокон. В соответствии с принципом изолированного проведения импульс проводится каждым волокном самостоятельно, без передачи соседним волокнам. Это не исключает возможности взаимодействия между параллельными волокнами. Показано, что местные токи, вызываемые деполяризацией в нервных волокнах, снижают порог в соседних волокнах на 20%. Но эти токи лежат ниже порога и не вызывают им- пульсации в волокне, на которое они действуют. Импульсы, бегущие по отдельным волокнам нервного пучка, могут быть ослаблены взаимодействием разных входных сигналов. Однако следует ясно понимать, что нервная система не могла бы функционировать без изоляции волокон.
Синхронное проведение. В центральной нервной системе изолированное проведение может сменяться синхронной активностью нервных волокон. Сильное взаимодействие между нервными волокнами, механизм которого еще не совсем понятен, может приводить к их одновременной деполяризации и в результате к синхронизированному проведению импульсов. Волокно, «диктующее» частоту импульсов остальным волокнам, действует как пейсмейкер (водитель ритма).
Диаметр аксона и скорость проведения. В более толстых волокнах перехваты отстоят дальше друг от друга, чем в более тонких волокнах. Поэтому в первых потенциал действия перемещается быстрее, так как ему приходится совершать более длинные скачки. Факим образом, существует тесная корреляция между диаметром волокна и скоростью распространения импульсов. Если известен диаметр аксона, можно вычислить
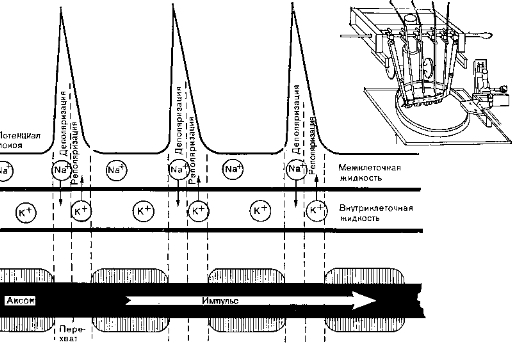
Рис. 4. Методика регистрации потенциала действия (вверху справа) и сальтаторное распространение импульсов. ПотеН' циал действия можно отводить только от перехватов Ранвье, которые не покрыты миелиновой оболочкой.
скорость проведения; и наоборот, если известна скорость проведения, из нее можно вывести диаметр аксона. Максимальная скорость проведения составляет 120 м/с, минимальная 1 — 2 м/с. По сравнению со скоростью распространения электрического тока даже самые крупные нервные волокна проводят импульсы очень медленно. Поэтому понятно, что время реакции, которое требуется для двигательного ответа и значительная часть которого идет на проведение, достигает нескольких миллисекунд.
Бинарные сигналы, т. е. серии импульсов, бегущие по афферентным нервам к центральным нейронам, однотипны. Сигналы, одновременно регистрируемые на многоканальном электроннолучевом осциллографе в нервах, идущих от зрительных, слуховых и осязательных рецепторов, не отличаются друг от друга. Эти совершенно неспециализированные сигналы расшифровываются нейронами центральной нервной системы. Двоичные сигналы декодируются, т. е. вновь преобразуются в аналоговые, в зонах центрального представительства сенсорных нервных путей.
Частотный код. Насколько мы теперь знаем, функция периферического рецепторного аппарата состоит в переводе стимулов внешнего мира в частотный код, пригодный для передачи. Именно это происходит, когда рецепторные потенциалы аналоговой природы, т. е. количественно зависимые от силы стимула, преобразуются в потенциалы действия одинаковой амплитуды, различающиеся только по частоте («всё или ничего»). Таким образом, частота спайков коррелирует с интенсивностью стимула. Этот механизм рецепторного кодирования, на который впервые указал Эдриан (Adrian), и поныне считается основой передачи информации в нервной системе. Однако это, как полагают, лишь один из элементов неизвестной системы кодов. В эту систему могут входить также пространственная интеграция импульсов, идущих по отдельным волокнам (которые могут оказывать друг на друга возбуждающее или тормозное действие), и взаимодействие между волокнами. Выяснение механизмов центрального декодирующего аппарата — одна из самых интригующих проблем психофизиологии.
Чтобы понять распространение коротких, периодических электрохимических импульсов, надо сначала обратиться к нейрону в состоянии покоя. Обычно разность потенциалов между внешней и внутренней сторонами мембраны рецепторного нейрона в покое составляет примерно от 30 до 100 мВ. Эту разность мы называем потенциалом покоя. Ее можно измерить, введя в клетку микроэлектрод толщиной около 1 мкм, а второй электрод поместив с внешней стороны клеточной мембраны. Разность потенциалов регистрируется достаточно чувствительным прибором. Пока еще не установлено, чем вызывается эта разность. Но одно можно сказать с уверенностью: внутриклеточная жидкость (жидкое содержимое клетки) по своему составу отличается от наружной, т. е. внеклеточной, жидкости. В то время как внутри клетки в избытке имеются ионы калия, в тканевой жидкости больше ионов натрия. Содержимое клетки богато белком, а в наружной жидкости много ионов хлора. Как полагают, высокую концентрацию калия и низкую концентрацию натрия в клетке создает работа так называемого «натриевого насоса» — особого активного метаболического процесса. Потенциал покоя, согласно Ходжкину (Hodgkin), достаточно хорошо объясняется неравномерным распределением ионов.
Аналоговый ответ. При раздражении рецепторного нейрона потенциал покоя снижается на 30—50 мВ, т. е. происходит частичная деполяризация. В таком состоянии достаточно второго стимула гораздо меньшей интенсивности, чтобы потенциал покоя исчез. Упомянутое снижение потенциала в рецепторе строго локально — оно происходит только в том участке клеточной
Потенциал поноя Рецепторный потенциал Потенциал действия
Рис. 1. Генерация и распространение нервного импульса в тельце Пачини и в аксоне, выходящем из рецептора. Rx и fe - положение регистрирующих микроэлектродов (внизу) и соответствующие записи (вверху): рецепторный потенциал и периодический потенциал действия.
мембраны, на который подействовал стимул. Еще одна особенность деполяризации состоит в том, что степень изменения потенциала зависит от интенсивности стимула: более слабые стимулы лишь ненамного снижают потенциал покоя, а сильные вызывают более выраженную деполяризацию. Можно сказать, что это локальное снижение потенциала пропорционально внешнему стимулу; поэтому в терминах кибернетики его можно назвать аналоговым ответом.
Такое локальное уменьшение разности потенциалов, носящее аналоговый характер, биофизики называют генераторным потенциалом. Генераторный потенциал рецептора некоторые авторы называют рецепторным потенциалом. Этот местный сдвиг потенциала представляет собой очень важный феномен, так как он порождает нервные импульсы в аксоне, связанном с рецепторной клеткой. Нервный импульс не остается локальным,
он распространяется и, как мы увидим, не является аналоговым, а носит — опять-таки на языке кибернетики — характер бинарного (двоичного) сигнала. Тельце Пачини. Рецепторный потенциал часто исследуют в тельцах Пачини, которые в большом числе имеются в коже и других тканях (например, в брюшине) человека и животных.
Рис. 2. Схема строения нейрона. Видно, как к верхнему нейрону подходят окончания аксона от другого нейрона. Короткие отростки—дендриты, длинные волокна с миелиновой оболочкой — аксоны (обратите внимание иа разрывы в оболочке, называемые перехватами Ранвье). Заштрихованные участки тела клетки изображают клеточную мембрану с сидящими на ней концевыми вздутиями. В остальной части клетки показаны ядро, ядрышко и гранулы тигроида. Эти последние играют важную роль в метаболизме и синтезе белка. Стрелками показано направление распространения импульсов.
Тельце Пачини по своей структуре напоминает луковицу; оно реагирует на давление, т. е. на механическую силу, изменяющую его структуру (рис. 1). В нервном окончании,погруженном в тельце Пачини, возникает рецепторный потенциал, после чего сигнал распространяется к другим частям нейрона в форме периодических импульсов.
Строение нейрона показано на рис. 2. В головном мозгу и остальной нервной ткани большинства животных тела нейронов имеют звездчатую или слегка округленную форму. Наиболее заметная особенность нейрона — наличие отростков, похожих на корешки. Короткие ветвящиеся отростки называются денд- ритамщ их функция обычно состоит в приеме сигнала. Длинный одиночный отросток, часто снабженный миелиновой оболочкой, называется аксоном. Функция его состоит в проведении нервных импульсов. В то время как некоторые нейроны лишены дендритов, ни одна нервная клетка ни у одного вида животных не может не иметь аксона. Аксой (или нервное волокно) — это самая важная для проведения импульсов структура, основной элемент нервных проводящих путей.
Бинарный ответ. Если рецепторный потенциал превысит порог возбуждения аксона, тотчас же произойдет его полная деполяризация. Это внезапное изменение называют потенциалом действия. Как мы видели, в рецепторе потенциал покоя только снижается. В аксоне же он совсем исчезает; более того, на одну тысячную секунды — или даже на меньший интервал — внутренняя часть волокна становится слегка электроположительной по отношению к окружающей жидкости.
Такая «реверсия» потенциала периодически повторяется. В момент максимума потенциала действия (спайка) мембрана аксона становится нечувствительной к последующим стимулам (рефрактерность), и новый импульс возникает лишь после того, как она восстановит свое нормальное состояние поляризации. Таким образом получается характерный для нервного проведения поток импульсов. Серия спайков распространяется вдоль нервного волокна, в миелинизированном волокне перескакивая с одного перехвата Ранвье (см. рис. 2) на другой. В отличие от местной деполяризации при возникновении импульсов деполяризация не варьирует с изменением интенсивности стимула. Будет ли пришедший сигнал пороговым или гораздо более сильным, амплитуда спайка остается одинаковой (ответ типа «всё или ничего»).
Два типа потенциала — два типа мембраны. Из сказанного выше видно, что генерация нервных импульсов в рецепторных структурах проходит две фазы. Первая фаза — это местная частичная деполяризация, создающая рецепторный потенциал. Когда это снижение потенциала достигает достаточной величины, деполяризуется также мембрана примыкающего аксона и
по нерву распространяется серия спайков (потенциалов действия). Различия в свойствах рецепторного потенциала и потенциала действия указывают на возможное различие свойств мембраны в соответствующих участках:
|
Мембрана аксона в рецепторе |
Мембрана аксона вне рецептора |
|
Электрическая реакция — частичная деполяризация Локальный (местный) процесс Корреляция между интенсивностью сигнала н степенью деполяризации (аналоговый ответ) Отсутствие рефрактерности |
Электрическая реакция — полная деполяризация Распространяющийся процесс Ответ типа «всё или ничего» (бинарный) Рефрактерное состояние после прохождения импульса |
Возникает вопрос: каким образом внешний стимул преобразуется рецептором в периодические импульсы? Или, говоря на языке кибернетики, каким образом в рецепторе происходит преобразование аналогового сигнала в бинарный? Как мы видели, рецепторный потенциал отражает интенсивность и длительность стимула: деполяризация пропорциональна силе входного сигнала, и она длится, пока длится стимуляция. Именно поэтому электрическую реакцию рецептора можно назвать аналоговым ответом, который отражает параметры стимула.
Место преобразования. Нервные сигналы типа «всё или ничего» мы находим при изучении электрических изменений в любой части периферического афферентного нервного пути. Отсюда можно сделать вывод, что преобразование аналоговых сигналов в бинарные, т. е. кодирование, совершается в рецепторе. Затем нервные сигналы передаются центральным нейронам в этой преобразованной форме, которая облегчает передачу, а возможно, и хранение (см. гл. II).
Информация. Под информацией обычно понимают сообщения или новые сведения о каком-нибудь событии или изменении. В этом смысле нервные импульсы, идущие от периферических рецепторов в головной мозг, передают информацию точно так же, как импульсы, бегущие по телефонному кабелю, или радиоволны. Чем меньше вероятность события, тем большую ценность представляет информация о нем. С помощью теории вероятности информацию можно измерить и придать ей численное выражение. Единицей при измерении количества информации, содержащейся в сообщении, служит бит. Это количество информации получается при единичном выборе типа «да или нет», если вероятность события равна 0,5. Теория информации интересуется только количественными сторонами информации, оставляя в стороне ее смысл и значение для получателя,
В отличие от этого нейрофизиология имеет дело также с качеством информации, т. е. с той пользой, которую организм может извлечь из полученного сообщения для своего благополучия и приспособления.
Кодирование. Сообщения передаются в нервной системе, как вообще в коммуникационных системах, в форме сигналов (кодов). Преобразование в определенную систему сигналов (кодирование) облегчает передачу и хранение информации. Необходимое условие состоит в том, чтобы сообщение могло быть декодировано в конечном воспринимающем звене. В случае нервной системы можно сказать, что кодирование происходит в рецепторах, а декодирование — в головном мозгу.
Бинарный код. Кодирующие системы, в которых используются только два вида сигналов (0 и 1), называются бинарными (двоичными)! В нервной системе информация, вероятно, передается с помощью такого бинарного кода. Во всяком случае, на это указывает правило «всё или ничего» для нервных сигналов, состоящих из серии импульсов, за которыми следуют реф* рактерные периоды.
Бинарные сигналы применяются для передачи и хранения информации также в компьютерах. Использование двоичной системы счисления, предложенное Нейманом (Neumann) и другими, удобно, так как позволяет применять простую и надежную электронную аппаратуру.
Информационный канал. Для передачи информации требуется соответствующая среда. Это то, что мы называем каналами (рис. 3). Информационными каналами служат, например, телефонные кабели и аксоны нервных клеток. Пропускную способность таких каналов определяют как максимальное количество информации, которое может быть передано в единицу времени; ее выражают числом битов в секунду. Сигналы, поступающие в систему извне, называются входными, а сигналы, выходящие на другом ее конце, — выходными. На практике часть информации при передаче теряется в результате искажения входных сигналов под влиянием различных факторов (так называемого шума), мешающих передаче сообщений в нервной системе. Вопрос о такого рода искажении, в частности в перехватах Ранвье по ходу миелинизированных нервных волокон и в других местах, составляет еще не решенную нейрофизиологическую проблему. Возможно, что искажения импульсации — бинарных сигналов, передаваемых по нервному волокну,— корректируются сигналами в других, параллельных волокнах, передающими ту же самую информацию. Как указывает Нейман, использование параллельных зашумленных каналов статистически дает благоприятные результаты. Таким образом, сравнительно слабая изоляция нервных волокон в конечном итоге улучшает функцию нервного пути посредством статистической
Закодированный Закодированный Первоначальный сигнал сигнал + шум сигнал
Рис. 3. Схема передачи информации в технических коммуникационных системах (вверху) и в нервной системе (внизу).
корреляции серий импульсов, идущих по нескольким каналам.
Кодирующий механизм рецепторов пока еще мало изучен. Два типа активности мембраны у аксона, связанного с рецепторной клеткой, указывают на то, что перекодирование должно происходить в месте соединения участков мембраны того и другого типа. В механорецепторах (например, в рецепторе давления — тельце Пачини) критической точкой является первый перехват Ранвье по ходу миелинизированного волокна, идущего от рецепторной клетки (см. рис. 1).
Сальтаторное проведение. Бинарный импульс, идущий по нервному волокну с миелиновой оболочкой, перескакивает от перехвата к перехвату. Такое сальтаторное проведение выявлено в эксперименте. Показано, что порог возникновения импульса в аксоне очень низок именно в перехватах Ранвье. Хаксли и Стемпфли (Huxley, Stampfli) отводили микроэлектродами потенциал действия от участка нерва между двумя перехватами и нашли, что на всем отрезке волокна между двумя перехватами потенциалы действия появляются одновременно без задержки в проведении. Такая задержка обнаруживается только между двумя соседними перехватами (рис. 4).
Изоляция нервных волокон. В соответствии с принципом изолированного проведения импульс проводится каждым волокном самостоятельно, без передачи соседним волокнам. Это не исключает возможности взаимодействия между параллельными волокнами. Показано, что местные токи, вызываемые деполяризацией в нервных волокнах, снижают порог в соседних волокнах на 20%. Но эти токи лежат ниже порога и не вызывают им- пульсации в волокне, на которое они действуют. Импульсы, бегущие по отдельным волокнам нервного пучка, могут быть ослаблены взаимодействием разных входных сигналов. Однако следует ясно понимать, что нервная система не могла бы функционировать без изоляции волокон.
Синхронное проведение. В центральной нервной системе изолированное проведение может сменяться синхронной активностью нервных волокон. Сильное взаимодействие между нервными волокнами, механизм которого еще не совсем понятен, может приводить к их одновременной деполяризации и в результате к синхронизированному проведению импульсов. Волокно, «диктующее» частоту импульсов остальным волокнам, действует как пейсмейкер (водитель ритма).
Диаметр аксона и скорость проведения. В более толстых волокнах перехваты отстоят дальше друг от друга, чем в более тонких волокнах. Поэтому в первых потенциал действия перемещается быстрее, так как ему приходится совершать более длинные скачки. Факим образом, существует тесная корреляция между диаметром волокна и скоростью распространения импульсов. Если известен диаметр аксона, можно вычислить
Рис. 4. Методика регистрации потенциала действия (вверху справа) и сальтаторное распространение импульсов. ПотеН' циал действия можно отводить только от перехватов Ранвье, которые не покрыты миелиновой оболочкой.
скорость проведения; и наоборот, если известна скорость проведения, из нее можно вывести диаметр аксона. Максимальная скорость проведения составляет 120 м/с, минимальная 1 — 2 м/с. По сравнению со скоростью распространения электрического тока даже самые крупные нервные волокна проводят импульсы очень медленно. Поэтому понятно, что время реакции, которое требуется для двигательного ответа и значительная часть которого идет на проведение, достигает нескольких миллисекунд.
Бинарные сигналы, т. е. серии импульсов, бегущие по афферентным нервам к центральным нейронам, однотипны. Сигналы, одновременно регистрируемые на многоканальном электроннолучевом осциллографе в нервах, идущих от зрительных, слуховых и осязательных рецепторов, не отличаются друг от друга. Эти совершенно неспециализированные сигналы расшифровываются нейронами центральной нервной системы. Двоичные сигналы декодируются, т. е. вновь преобразуются в аналоговые, в зонах центрального представительства сенсорных нервных путей.
Частотный код. Насколько мы теперь знаем, функция периферического рецепторного аппарата состоит в переводе стимулов внешнего мира в частотный код, пригодный для передачи. Именно это происходит, когда рецепторные потенциалы аналоговой природы, т. е. количественно зависимые от силы стимула, преобразуются в потенциалы действия одинаковой амплитуды, различающиеся только по частоте («всё или ничего»). Таким образом, частота спайков коррелирует с интенсивностью стимула. Этот механизм рецепторного кодирования, на который впервые указал Эдриан (Adrian), и поныне считается основой передачи информации в нервной системе. Однако это, как полагают, лишь один из элементов неизвестной системы кодов. В эту систему могут входить также пространственная интеграция импульсов, идущих по отдельным волокнам (которые могут оказывать друг на друга возбуждающее или тормозное действие), и взаимодействие между волокнами. Выяснение механизмов центрального декодирующего аппарата — одна из самых интригующих проблем психофизиологии.
Источник: Адам Д., «Восприятие, сознание, память. Размышления биолога» 1983