Импульсная активность нейронных популяций при мыслительной деятельности
Первый этап исследования. Мы имели возможность находить точки мозга, более или менее тесно связанные с психической деятельностью. Для расшифровки протекающих в них процессов физиологу предстояла сначала задача выбрать из регистрируемых биоэлектрических процессов - наиболее подходящий для этой цели. Первым (хотя и не единственным) кандидатом на данную роль была импульсная активность нейронов, прежде всего, в связи с быстродействием мыслительных процессов. Далее встал вопрос: отдельная нервная клетка или функционально объединенное сообщество нервных клеток? Многолетний опыт экспериментальных исследований (Василевский, 1968; Альтман, 1972, и др.) показал, что одна нервная клетка чаще всего не способна отражать своей активностью сложные сигналы. При регистрации динамики отдельной или, точнее, отдельных клеток как представительных единиц в человеческом мозгу можно получить ряд интересных сведений о его работе (см. выше). Однако на уровне группы нервных клеток, работающих в том нужном согласии друг с другом, которое создает новое качество, можно было надеяться найти отражение уже более сложных сигналов (Коган, 1973). Вопросы о свойствах популяции нервных клеток подробно рассматриваются Буллоком (Bullok, 1979).
Таким образом, еще в период, когда сравнительно редко использовалось отведение импульсной активности не с микроэлектродов, а с так называемых микромакроэлектродов (в нашем случае с диаметром 50-100 мкм и рабочей поверхностью от 0.01 до 0.15 мм ), было начато изучение реорганизации импульсной активности нейронных ансамблей при простейших видах мыслительной деятельности. С вживленных в различные структуры мозга (кору, образования таламуса и стриопаллидарной системы, верхние отделы ствола, медиобазальные отделы височных долей) электродов регистрировалась активность популяций нервных клеток. В этой активности обычно обнаруживались пики разрядов нескольких нервных клеток, более или менее существенно превышавшие по амплитуде труднодифференцируемую активность большого количества нервных клеток. Варианты регистрируемой с вживленных электродов активности представлены на рис. 14.
Любое исследование тонких нейрофизиологических коррелятов мыслительной деятельности до последнего времени имело и еще, по-видимому, в течение ряда лет будет иметь, кроме собственно нейрофизиологических, по крайней мере два аспекта. Так, во-первых, в каждом из них в соответствии с нейрофизиологическими задачами требовалось найти адекватный психологический подход. И хотя многие из применявшихся тестов строились на основе классических психологических проб, далеко не все могло быть использовано в традиционном варианте (тест Вине и др.). Во-вторых, возможности нейрофизиологического анализа ответов активной, реагирующей на психологический тест точки существенно зависели от степени совершенства математико-технической обработки извлечения полезного сигнала.
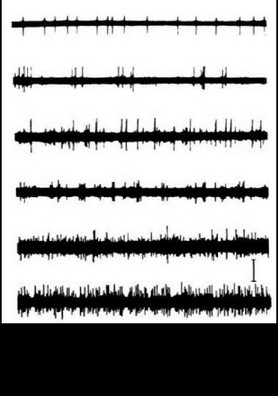
Сверху вниз:
поясная извилина, премоторная область коры, вентролатеральное таламическое ядро срединный центр, заднее вентральное латеральное таламическое ядро, бледный шар. Калибровка: 20 мс, 50 мкВ
Для избежания разночтений уточним, в чем разница между популяцией нервных клеток и их рабочим ансамблем. С электрода регистрируется активность группы нервных клеток. Это - всегда популяция, но не всегда ансамбль. Как можно проверить подобное утверждение? Дается задача, одна, другая - сосчитать что-то в уме, повторить ряд слов. Затем с помощью специальных программ и ЭВМ изучается реакция этой популяции нервных клеток. Какие здесь возможны варианты? Может просто не произойти ничего существенного - и тогда, скорее всего, данная популяция не имеет отношения к реализуемой деятельности. Реакция может быть относительно простой - в виде общего тонического увеличения или уменьшения частоты разрядов.
В этом случае следует предположить, что данная популяция вносит в систему обеспечения психической деятельности «энергетический» вклад того или другого знака или что существующие методы еще не способны обнаружить более тонкие изменения. Может выявиться, наоборот, перестройка организации работы нервных клеток. И тогда правомерно использовать термин «ансамбль», хотя в конкретном
случае это может быть лишь часть ансамбля, так как остальная находится за пределами возможностей регистрации активности ее клеток с данного электрода. А может быть и иначе. Только часть популяции существенно реорганизуется, работает в данных условиях как слаженный ансамбль. Но вот задается другая деятельность, ансамбль захватывает другие клетки из популяции или, наоборот, становится меньше. Ансамбли очень динамичны, они все время «дышат». Наверное, тем, кто не работает непосредственно в данной области, представить себе организацию и реорганизацию ансамблей можно, вспомнив динамичную световую рекламу, - из набора однотипных источников света слагаются ежемоментно меняющиеся, иногда очень сложные световые узоры. Понятно, что не только это сравнение, но и приведенные выше положения - лишь схема, которую иногда очень нелегко уточнить в процессе математико-технической обработки.
Те зоны, в которых нами обнаруживались воспроизводимые изменения физиологических показателей мозга при психологических пробах, не относились к анатомическим образованиям акустического (слухового) анализатора. Лишь теперь, через много лет после начала наших исследований, у тех, кто работает в области мозгового анализа звуковых сигналов, начал пробуждаться интерес именно к этим зонам, в которых, как предполагают Эммере (Emmers, 1970) и Я. А. Альтман с соавторами (1979), происходит обработка сложных сигналов, высшая по отношению к протекающей в классических образованиях анализатора. И, как считает Б. Ф.
Толкунов (1978), именно в этих зонах существуют оптимальные анатомофизиологические условия для высшей интеграции. Тем не менее самым началом нашего углубленного изучения тонких нейрофизиологических коррелятов психической деятельности были поиск и отражения акустических характеристик слов и их элементов в активности популяций (ансамблей) нейронов структур стриопаллидарной системы, таламуса, медиобазальных отделов височных долей, верхних отделов ретикулярной формации не как дань представлениям Пенфильда (Penfield, 1975) о центроэнцефальной системе, а как результат собственного опыта. Как указывалось, именно в этих зонах отмечались воспроизводимые реакции при психологических пробах. При этом мы отдавали себе отчет в том, что поиск коррелятов может быть затруднен непрямым отражением акустических свойств сигнала в связи с характером образований.
В результате первых исследований, по ходу которых проводилась весьма трудоемкая отработка методов анализа мультиклеточной активности, была показана, хотя и непрямая, связь развивающихся в области этих подкорковых структур пространственно-временных перестроек импульсной ак тивности с частотноамплитудными характеристиками входного сложного звукового сигнала - слова (Бехтерева и др., 1971). Известную роль в прогрессе исследований сыграло предложенное на основе работ Сондерсона с соавторами (Sanderson et al., 1973) и П.
В. Бундзена (1976) использование так называемой дистрибутивной формы
представления пространственно-временны^х перестроек импульсной активности нейронных популяций. Эта форма не только открыла новые возможности машинного анализа импульсной активности при психологических пробах, но и явилась необходимым методическим толчком для изучения значимых элементов в перестройках мультиклеточной активности. Детали математико-технической обработки импульсной активности приведены в работах П. В. Бундзена с соавторами (19756), Ю. Л. Гоголицына, П. Д. Перепелкина (1975), Ю. Д. Кропотова (1976, 1978), П. Д. Перепелкина (1977, 1978), в разделе П. Д. Перепелкина в работе Н. П. Бехтеревой с соавторами (1977а). Идет постоянное совершенствование методов на основе общей принципиальной схемы анализа. По ходу изложения материалов там, где без этого нельзя обойтись, будет приведена физиологическая логика этого математико-технического анализа.
Как видно из наших первых работ, слово как сложный акустический сигнал и - что в этих работах не исключается, но и не доказывается, - как сигнал семантически значимый вызывает тонические и физические перестройки импульсной активности. Одновременно в данных работах показано, что упомянутые перестройки обнаруживаются только в части популяций, а ансамблем может являться часть большой популяции. Как это обнаруживается? Если при работе с отдельными нервными клетками для получения представления о зоне мозга исследователи идут неизбежно по пути накопления данных о поведении нейронов в этой зоне, то при работе с популяциями нервных клеток методической основой последующей интегративной фазы, объединения, является первоначальное разделение популяций на достаточно малые группы нейронов. Осуществляется выделение тех разрядов нейронов, которые, превышая уровень физиологического шума, позволяли проводить исследования при минимально возможных искажениях мозговых сигналов - имеется в виду прежде всего искажение формы разряда при регистрации в результате суперпозиции двух и более спайков. С этой целью мы прежде всего разделяли разряды по амплитуде. Такой метод при некоторой его уязвимости и используется не только у нас, ной в ряде других исследований (Keidel, 1975; Schwartz et al., 1976; Halgren et al., 1978; Ravagnati et al., 1979). Разделение по амплитуде производится в виде так называемой «оконной» дискриминации (Бехтерева и др., 1973), предполагающей выделение нескольких амплитудных уровней разрядов. При этом в каждое «окно» попадают именно те разряды, пик которых оканчивается в его пределах. Именно на основе этой оконной дискриминации и были разделены разряды мультиклеточной активности в наших первых исследованиях.
Наравне с «оконной» дискриминацией в связи с накоплением достаточного материала и отсюда возможности выбора сейчас предпочтительно используется определенный амплитудный порог, зависящий от соотношения амплитуд выделяемых отдельных разрядов и физиологического шума более низкой амплитуды, «щетки», искажающих друг друга разрядов многих клеток; обычно этот порог определяется в размере 0.7 от максимальной амплитуды разрядов. При этом, естественно, учитывается лишь часть регистрируемой активности, но, что очень важно, ее наименее искаженная или даже практически неискаженная часть.
В обоих случаях обычно оказывается возможным обнаружить большее или меньшее количество разрядов равной и разной амплитуды. При исследовании активности отдельных нейронов амплитудный показатель может до известной степени и на небольшом промежутке времени помочь идентифицировать, опознать нейрон. Сходно обстоит дело и при дифференциации мультиклеточной активности. Однако идентификация отдельных нейронов более надежна, хотя тоже не абсолютна, при использовании и амплитудного, и дополнительного показателя - формы потенциалов действия. Этот прием также с 1973 года в течение ряда лет использовался в нашей лаборатории.
При регистрации мультиклеточной активности оказывается возможным выделить с небольшими искажениями динамику активности до четырех, редко более, отдельных нейронов, то есть принципиально то же, что возможно и в условиях аналогичного собственно микроэлектродного исследования.
Существенный выигрыш за счет регистрации именно мультиклеточной активности, а не активности отдельных нейронов, связан с возможностью изучения в таких условиях взаимодействия не только удаленных нейронов, но и нейронов, расположенных заведомо в пределах рабочего ансамбля. В эксперименте этот вопрос мог бы быть решенным за счет введения ряда микроэлектродов с заданным и фиксированным расположением между ними, однако до последнего времени такой прием в клинических исследованиях является неоправданным, несмотря на работы в области зрительного и слухового протезирования (Simmons et al., 1965; Brindley, Lewin, 1968; Brindley, 1970; Michelson, 1971). He исключено также, что для нейрофизиологического анализа это не представит особых преимуществ, хотя данный вопрос лучше оставить пока открытым. Проблема чистоты получаемых данных решается нами иначе - применением в исследованиях мозга человека электродов, у которых возможно произвольно варьировать рабочую поверхность и, таким образом, одновременно или последовательно записывать большее или меньшее количество нейронов (Матвеев, Киселев, 1978). Взаимодействие нейронов в пределах одной зоны мозга и между разными его зонами определялось первоначально путем сравнения активности, ее текущей частоты предварительно дискриминированных (способом оконного различения и др.) нейронов, причем основным приемом служил факторный анализ, а эпоха анализа избиралась в соответствии с задачами и плотностью выделенной активности.
Далее с этой целью применялись и другие приемы, и прежде всего изучение регулярности и связности импульсного потока в форме разных видов интервального анализа и прием когерентной детекции - выявления синхронности событий в удаленных зонах мозга (Бундзен идр., 19756; Кропотов, 1976, 1978; Перепелкин,
1977; Бундзен, Давид, 1979). Эти исследования проводились с целью уточнения не только взаимодействия между нервными элементами, но и всех перестроек местной активности нейронных популяций, характерных для исследуемого вида деятельности. Стимулирующими факторами данного направления работ служили следующие.
- Экспериментальные исследования, в которых описывалась регулярность и связность импульсных потоков в активности отдельных нейронов и нейронных популяций и возможность развития микроструктурных особенностей паттернов межимпульсных интервалов - узоров межимпульсных разрядов (Gerstein, 1960; Poggio, Vernstein, 1964; Kiang, 1965; Nakahama, 1966; Perkel et al., 1967;. Perkel, Bullok, 1968; Segundo, Perkel, 1969; Verzeano et al., 1970; Василевский и др., 1972; Babb et al., 1973; Василевский, 1976; Nakahama et al., 1977; Толкунов, 1978; Чораян, 1978; Шульгина, 1978, и многие другие).
- Специальное изучение чисто частотного кодирования, проведенное Эхорном с соавторами (Echorn et al., 1976), показавшее, что при этом виде кодирования возможные потери информации могут быть очень велики - от 10 до 95 %!
- Данные о функциональных и морфологических основах возможности кодирования появлением специфических узоров импульсной активности (Anderson et al., 1964; Schlag, Villabanca, 1968; Gerstein, Perkel, 1969; Emmers, 1970, 1976; Moore et al., 1970; Sanderson et al., 1973; Katayama, Murata, 1974; Baldissera, Parmiggiani, 1976; Толкунов, 1978).
- Известная закономерность об усилении роли кодов в процессе эволюционного развития и становления высших форм деятельности (Ашмарин, 1975; Николаев, 1976; косвенно данные: Богданов, 1978).
Уже в самом начале изучения взаимодействия нейронов было обнаружено, что в процессе психической деятельности наряду с изменением текущей частоты разрядов происходят и другие характерные изменения импульсной активности нейронных популяций. Ее местная перестройка в исследуемых структурах при психической деятельности характеризовалась не только изменением частоты, но и появлением удерживающейся в течение секунд, десятков секунд и - реже - минут мозаики и динамики пространственных взаимоотношений между близлежащими и удаленными нейронами в дистантно расположенных зонах мозга, что выражалось, в частности, появлением определенной структуры разрядов нейронной популяции - их определенного паттерна.
Именно поэтому в физиологическом плане вначале для, скажем так, проверки «проходимости пути» выгодной оказалась дистрибутивная форма представления развивающейся при исследуемой деятельности перестройки паттерна, так как она сохраняла информацию о пространственно-временны^х отношениях нейронов и
оставляла возможность их последующего, более углубленного исследования.
Дистрибутивная форма представления паттерна отражала динамику межимпульсных интервалов между разрядами группы нейронов, причем количество исследуемых (записываемых) нейронов определялось уровнем амплитудного среза. В плане возможностей исследования мультиклеточной активности с помощью ЭВМ при применении эталонного поиска паттерн в дистрибутивной форме в виде конечной (ограниченной временем) численной последовательности длительностей интервалов представлял собой приемлемый эталон, который мог быть заложен в ЭВМ (рис. 15) (Перепелкин, 1977). У каждого больного для каждой нейронной популяции в определенные отрезки времени накапливались дистрибутивные представления паттернов мультиклеточной активности при восприятии, удержании в памяти и воспроизведении различных слов (Бундзен и др., 19756). Так формировался банк эталонов, позволяющий у того же больного в соответствующих нейронных популяциях и обычно в связи с кратковременностью жизни этих паттернов в той же записи обнаруживать момент появления соответствующих паттернов и таким образом определять возникновение «слов» в мозгу не только в соответствии с непосредственным содержанием входного и выходного сигналов - предъявляемых слов или изображений и ответно произносимых слов, но и того, что имело к этим сигналам смысловое косвенное отношение. Следует подчеркнуть, что в связи с указанными выше временными ограничениями работы с эталонным поиском целесообразнее всего проводить (что и проводилось далее) в реальном времени исследования.
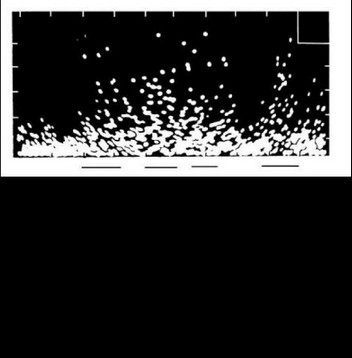
Вслед за использованием дистрибутивного представления паттерна начался поиск его значимых элементарных составляющих.
Подчеркнуты моменты воспроизведения различных цифр
В основу был положен тот же принцип - обнаружение связностей, групповых последовательностей разрядов с определенными соотношениями интервалов. Важность задачи поиска элементарных составляющих пространственно-временно^й реорганизации мультиклеточной активности, развивающейся при восприятии, удержании в памяти и воспроизведении слов, определялась рядом положений.
Прежде всего дистрибутивное представление паттерна, полученное на основе оконной дискриминации или амплитудного среза, не обязательно содержало только полезный сигнал. Возможность использования паттерна в эталонном поиске для обнаружения с помощью ЭВМ аналогов в мультиклеточной активности показывала значимость его как выражения тонких нейрофизиологических коррелятов слов. Но ни этот прием, ни какой-либо другой из применявшихся ранее, подтверждающие значимость фактора длительности межимпульсных интервалов в паттерне, не свидетельствовали о чистоте данного представления паттерна, не давали сведений о соотношении в нем сигнала и шума.
Эталонный поиск использовался в физиологии и ранее. Так, например, без особой необходимости для диагностики, но как важный элемент последующего управления биоэлектрической активностью больных эпилепсией эталонный поиск с помощью ЭВМ применялся уже в 60-х годах при использовании в качестве эталона эпилептиформной активности комплекса пик—волна. В этом случае, как вполне понятно, феномен спайк—волна мог быть выделен и при использовании менее информативного эталона - только спайка или только медленной волны. Но тогда наравне с сочетанным биоэлектрическим феноменом на ЭЭГ неизменно выделялись бы (распознавались!) и отдельные, и групповые спайки, и различные медленные волны. Использование дистрибутивного представления паттерна мультиклеточной активности принципиально могло быть таким же. Дистрибутивный эталон мог также представлять собой как бы часть сигнала. Образно (а может быть, и не очень образно, а реально?) он отражал бы часть нейрофизиологических коррелятов слова - его слог, триграмму, встречающиеся и в других словах. С другой стороны, в дистрибутивном представлении в зависимости от ряда условий и прежде всего просто от уровня амплитудного среза импульсной активности может содержаться и избыточная информация, загрязняющая эталон и, как это ни парадоксально, другим путем ведущая к тому же результату - возможности опознания с его помощью не только тех лов, на основе нейрофизиологического отражения которых он был образован.
Сложность вопроса усугублялась тем, что паттерн мог обнаруживаться в мозгу и в полном, развернутом виде, по времени соответствующем слышимому или произносимому слову, и - особенно фазу удержания слов в памяти или считывания из памяти - в сжатой, компрессированной форме. Действительно, уже в самых первых исследованиях в нашей лаборатории было показано, что после восприятия слова непосредственно за развернутым, полным паттерном в мозгу появляется компрессированный паттерн, содержащий опорные элементы первого.
Этим не кончаются сложности (хотя их не следует и переоценивать!) работы с «неочищенным» паттерном и любой формой его представления. Так, например, при эталонном поиске наиболее адекватным оказалось использование дистрибутивного представления не только той перестройки импульсной активности, которая соответствует по времени моменту произнесения слова больным, но и несколько большего ее отрезка, увеличенного за счет перестройки, предшествующей ответу, за счет захвата в эталон и фазы формирования ответа. Значимость этой фазы для ответного речевого сигнала подтверждается в ряде работ и прямо, в частности, в исследованиях С. Н. Раевой (1972, 1977), Хелгрина с соавторами (Halgren et al., 1978), и косвенно - в работах другого направления, связанных с регистрацией состояний мышц при произнесении слов (Syka, Schramm, 1975).
Все эти и ряд других соображений определили работу по выделению более элементарных составляющих паттерна.
Если первоначально за основу был взят паттерн, соотносимый со словом или его элементом, то далее методически и технически задача решалась самыми разными способами, в том числе изучением фоновой активности и активности при выполнении психологических тестов с соотнесением данных с фоновыми и тестовыми не только a priori, нон a posteriori проведенной обработки.
Математико-технический аспект данных направлений исследования мультиклеточной активности был описан в 1977 году. Здесь важно подчеркнуть, что в результате проведенных работ (Бехтерева, Бундзен, 19746; Гоголицын, 1974; Перепелкин, 1974; Бундзен, 1976, 1977; Малышев, 1977; Шкурина, 1978) в отрезках записи мультиклеточной активности продолжительностью обычно в несколько десятков секунд были выделены групповые последовательности разрядов с фиксированным ( +10 %) интервалами, причем функционально значимым элементом общего паттерна оказывались последовательности из трех и более интервалов. При этом, хотя и наблюдались групповые последовательности разрядов с определенными интервалами в количестве более 3 (до 6-7), они были менее стабильны, уникальны и менее воспроизводимы. Это позволило считать последовательность из трех интервалов минимальной и достаточной для физиологической роли ее как элемента паттерна. Чем характеризовалась функциональная значимость групповой последовательности разрядов с определенными интервалами? Первым опорным критерием было появление или существенное учащение этих триад в период психологического теста, соотносимое с его фазами, воспроизводимое у одного того же больного в аналогичных условиях. Вторым критерием была возможность использования групповой последовательности разрядов в качестве образца при эталонном поиске. Иными словами, если использование какой-то данной триады в качестве эталона позволяло выявлять в мультиклеточной активности моменты появления в мозгу общих паттернов, в которых присутствовала данная триада, она расценивалась как значимый элемент паттерна. Не перебирая все критерии, следует подчеркнуть еще один, не относящийся к конкретной триаде или квадриаде ит. п., но характеризующий значение этих групповых последовательностей как элементов паттерна, являющегося тонким нейрофизиологическим коррелятом психических процессов. Это - показанная В. Н. Малышевым (1979) возможность встраивания триад в паттерн при введении квазислова в смысловое поле определенных слов. Естественно, при этом удовлетворялись и первые два критерия.
Что представляет собой фоновая, или, точнее, так называемая фоновая, активность с точки зрения возможности определения в ней групповых последовательностей разрядов с определенными интервалами? Можно ли обнаружить в фоновой активности те же триады? Нередко можно, но обычно с меньшей вероятностью (Перепелкин, 1978). Что действительно обнаруживается в фоне в большом количестве, так это парные интервалы с определенной их длительностью, в том числе и одинаковой в обоих интервалах. Именно парные интервальные последовательности, по-видимому, служат физиологической основой образованийтриад (Малышев, 1977), а методически являются основой одного из наиболее перспективных приемов их поиска (Бундзен, 1977). Обнаружив парные интервалы, дальше с помощью соответствующих программ мы исследовали, не «привязывается» ли к ним в связи с тестом и не удерживается ли до или после этой пары еще один интервал, образуя таким образом триаду. По-видимому, парные интервалы могут рассматриваться в качестве опорных жестких единств для формирования интервальных триад. Однако для понимания механизмов формирования значимых групповых последовательностей разрядов с тремя и более интервалами определенной продолжительности нуждается в дальнейшем уточнении вопрос о парных интервальных последовательностях - являются ли они результатом развертывающихся генетических свойств мозга, процесса обучения в общем виде или обучения более конкретного, речевого.
Одна триада на определенном отрезке времени исследования может быть функциональным элементом нейрофизиологического паттерна и служить эталоном для машинного исследования импульсной активности одного или нескольких близких по смыслу слов. Однако это не общее правило, единственной триадой паттерн не исчерпывается даже в одной нейронной популяции. Каждый паттерн состоит из совокупности групповых последовательностей разрядов, и, таким образом, отражение слова или его элемента в импульсной активности нейронных популяций характеризуется набором этих групповых последовательностей разрядов. Говоря об интервальных последовательностях, следует помнить, что функционально значимым фактором является пространственно-временна^я перестройка межнейронных отношений, а они - ее отражением. При этом в разных, близких и удаленных друг от друга, зонах мозга одно и то же слово или его элемент, как правило, вызывает появление паттернов, состоящих из групповых последовательностей разрядов, главным образом своих для каждой зоны мозга, но также и очень сходных для разных зон (Бехтерева и др., 1977а). В 1980 году нами высказывалось предположение, что сходные групповые последовательности разрядов являются своего рода объединяющим, системообразующим фактором. Эти предположения нашли прямое подтверждение в исследованиях Г. А. Вартаняна и А. А. Пирогова (1987). Что же касается разных групповых последовательностей разрядов в разных зонах мозга, то они, повидимому, характеризуют прежде всего распределенность паттерна - распределенность кодирования психических процессов (Бехтерева, 1977). Исследование общих элементарных единиц и синхронных разрядов в разных нейронных популяциях, а также в разных элементарных единицах является задачей дальнейших исследований. Предполагается, что результаты этих исследований могут пролить свет на вопрос о том, какое новое качество приобретает популяция по сравнению с отдельной нервной клеткой, несколько звеньев системы - по сравнению с одним звеном, система - по сравнению со звеном или совокупностью звеньев. Надо подчеркнуть, что этот вопрос ставился нами с первых шагов данного исследования (Бехтерева, 1966, 1971).
При прослеживании динамики синхронных разрядов в работающих при тесте на принятие решения нейронных популяциях видно, что процесс принятия решения характеризуется динамическим увеличением синхронизации - сходства в разных зонах мозга. Есть известные основания полагать, что дальнейшее изучение проблемы приведет к обнаружению в паттернах и на уровне системы, и на уровне нейронных популяций жестких и гибких элементов (более или менее устойчивых). Это не противоречит основному положению о распределенности перестроек, но подтверждает общность принципа жестких и гибких элементов на разных уровнях организации мозга.
Чем дольше продолжалось изучение вопроса, тем больше подтверждалось, что ансамбль по сравнению с суммой клеток обладает дополнительными свойствами. При исследовании амплитуды и формы спайков оказалось, что групповые последовательности разрядов с определенными интервалами, и в частности триады, образуются (могут образовываться!) за счет разных нейронов, а не одного и того же нейрона. Таким образом, принцип пространственно-временно^й организации перестроек импульсной активности обнаружился и на уровне элемента паттерна.
На сегодня это можно объяснить с помощью следующей гипотезы: ансамблем нейронов является функциональное единство из ряда групп нейронов, каждая из которых обладает своими свойствами. При изменении состояния в активной зоне мозга под влиянием тех или иных факторов (приходящий к нейронам сигнал, изменение уровня базисных медленных электрических процессов ит. д.) срабатывает один нейрон из группы сходных нейронов, причем другие сходные нейроны в этом случае тормозятся, а нейрон из другой группы разряжается через временной интервал за счет прежде всего синаптической задержки при не всегда прямой передаче возбуждения (разного рода прямые и обратные связи). При этом синаптическая задержка играет роль квантующего фактора. Естественно, возможны и другие, также гипотетичные, варианты, в частности те, которые обсуждаются в работах Эммерса (Emmers, 1970) и были упомянуты нами выше. Морфологические данные (Поляков, 1973) о наличии разных нейронов и глиальных клеток даже в небольших клеточных скоплениях мозга как будто бы не противоречат такой точке зрения. Но, естественно, этот вопрос нуждается в дальнейшем изучении. Вопрос все тот же - часть и целое, нейрон и ансамбль и т. д.
При обсуждении методологических проблем современной нейрофизиологии уже говорилось о том, что далеко не все применяемые в психологии методы могут быть использованы в комплексном исследовании при изучении нейрофизиологических коррелятов психической деятельности. Психологический аспект исследования должен соответствовать нейрофизиологическим задачам. При этом на разных этапах исследования требования к психологической его части различны.
На том этапе, когда задачей нейрофизиологического исследования становится выделение первоначально общих перестроек импульсной активности в связи с конкретными словами, а затем все более тонких элементов этих перестроек, тесты существенно меняются. Так, например, произносятся одиночные синтетические гласные фонемы - ответ больного заключается в воспроизведении этой фонемы или того, что по его мнению, было ему предъявлено (случай предъявления так называемой смешанной синтетической фонемы, которая могла бы быть распознана и как [А], и как [Е]). Произносятся слоги или триграммы, причем их немного, нередко - один слог или одна триграмма, больной повторяет их в зависимости от задания сразу же или по команде. Называются отдельные слова или маленькие группы слов - знакомых, часто или редко встречающихся в речи, квазислова, слова на незнакомых больному иностранных языках; больной повторяет их также непосредственно после предъявления или по команде. Все это достаточно просто для больного, и какая-либо собственно психологическая сложность задачи при этом отсутствует. Однако, как вполне понятно, именно этот этап исследования исключительно важен не только для нейрофизиологии, но и для психологии как науки. Только при сочетанном психонейрофизиологическом исследовании может быть получен надежный ответ на вопрос, что реально происходит в мозгу при восприятии слова, в момент, когда формируется оперативная единица психической деятельности, в момент подготовки и реализации вербального ответа и т. д. Существующие представления о наличии в мозгу возникающей в процессе обучения «фонемной решетки» (Жинкин, 1958) интересны, но, вероятно, не могут претендовать на абсолютную универсальность.
Рабочей гипотезой в проводимых исследованиях были представления о том, что слово может быть отражено в мозгу пофонемно, в виде отдельных слогов и больших блоков, особенно при частом употреблении его в речи. При этом принцип пофонемного кодирования остается, по-видимому, неизменным при обучении и
вербализации ответа. Однако, чтобы использовать получаемые нейрофизиологические данные для понимания механизмов собственно мыслительной деятельности, необходимыми этапами были исследование отражения в выделяемом импульсном паттерне смысловых свойств слова и далее построение и предъявление задач, позволяющих судить о нейрофизиологических основах собственно мыслительной деятельности человека. Зоны, участвующие в обеспечении психической деятельности, достаточно достоверно можно было обнаружить вначале с помощью метода постстимульныхгистограмм (Гоголицын, 1979; Кропотов, 19766), ав последующие годы - перистимульных гистограмм (ПСГ) (Гоголицын и др., 1987а).
В этом случае удается обнаружить (например, в передних отделах поясной извилины) и некоторую разницу реакций на слова разных смысловых полей (рис. 16). Однако вполне понятно, что прямое решение этой задачи если не невозможно, то во всяком случае весьма сложно, а поэтому изучение отражения смысловых свойств в перестройках импульсной активности нейронных популяций было предпринято сотрудниками нашей лаборатории в форме поисков признаков смысловой общности или, что в данном случае принципиально то же самое, смысловой дифференциации на основе модификации теста Озгуда (Osgood, 1953).
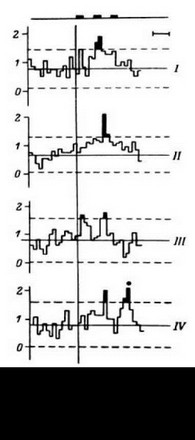 ПСГ активности нейронной популяции
ПСГ активности нейронной популяции
- 3 нейрона. Каждая гистограмма
накоплена по 16 предъявлениям пар слов, которые обобщались по смыслу пациентом. По оси абсцисс - время (калибровка - 1 с, бин гистограммы - 200 мс). Зачерненные прямоугольники вверху - средние эпохи звучания двух предъявляемых слов и ответа пациента. Вертикальная линия - начало теста. По оси ординат - среднее число разрядов нейронов в бине гистограммы за одно предъявление. Штриховыми линиями ограничен доверительный интервал, соответствующий 99%-ной вероятности отклонения частоты импульсации от уровня фона. I - гистограмма, накопленная при предъявлении пар слов, обозначающих предметы мебели; II - то же - посуды; III - то же - названий деревьев; IV - смешанная группа (пары слов не могли быть обобщены и пациент отвечал «нет»). Черный кружок - дополнительный всплеск активности, возможно, эмоциогенный. Заметно также большее возрастание частоты разрядов при предъявлении второго слова
Больному назывались последовательно слова, которые могли бы быть обобщены по смыслу каким-то другим понятием. Так, например, перечислялись предметы мебели: «стул», «стол», «шкаф» и т. д., после чего больного просили назвать обобщающее слово, как вполне понятно, в данном случае - «мебель». Задача могла быть поставлена и в еще более облегченной для больного форме - ее условия формулировались заранее: «Вам будет назван ряд слов. Постарайтесь уловить в них общий смысл и назвать обобщающее понятие». Если в тестах перечислялись названия цветов, больной отвечал: «Цветы», если деревьев, - ответ был: «Деревья».
Далее предпринималась задача встраивания нового слова в смысловое поле, сформировавшееся у больного в онтогенезе на основе биосоциального опыта (Малышев, 1979). Так, например, попеременно перечислялись названия цветов и деревьев и в ходе теста (обычно не в начале ннев конце) без предупреждения о том, что в смысловом поле будет предъявлен не имеющий смысла элемент, произносилось квазислово. Если больной спрашивал, что это такое, в ходе теста ему отвечали, что это - название цветка или дерева. В последующем больному сообщалось, что данный словоподобный сигнал значения не имеет. При изучении нейрофизиологического отражения смысла слов применялись пробы, где одно и то же по звучанию слово имело в зависимости от конте
Источник: Наталья Петровна Бехтерева, «Здоровый и больной мозг человека» 2010
А так же в разделе «Импульсная активность нейронных популяций при мыслительной деятельности »
- Введение
- Глава первая Современные принципы и возможности изучения мозга человека
- Глава вторая Некоторые общие принципы организации мозга человека
- Артифициальные стабильные функциональные связи — новый феномен долговременной памяти
- О гибких и жестких звеньях мозговых систем обеспечения психической деятельности
- Глава третья Общие механизмы мозга и болезнь
- Устойчивое патологическое состояние
- Воздействия на устойчивое патологическое состояние
- Защитные механизмы мозга
- Глава четвертая Нейрофизиология эмоций [3 - В данной главе использованы материалы совместных работ и публикаций с В. М. Смирновым и Д. К. Камбаровой.]
- Глава пятая Нейрофизиология мышления
- Глава шестая Нейрофизиология памяти человека и общие механизмы его мозга
- Глава седьмая Ближайшие перспективы в физиологии мозга человека на основе ее сегодняшних возможностей
- Литература
- О мозге человека. XX век и его последняя декада в науке о мозге человека
- Некоторые положения в развитии проблемы. Факты и концепции
- Перспективы развития проблемы (возможен ли третий прорыв?)
- Жорес Иванович Алферов (вступительное слово)