Мышечные ткани
Мышечные ткани включают исчерченную (поперечнополосатую), неисчерченную (гладкую) и сердечную. Эти разновидности мышечной ткани имеют различное происхождение и строение. Мышечные ткани объединены по своему строению и по функциональному признаку — способности сокращаться, изменять свою длину, укорачиваться.
Исчерченная (поперечнополосатая, скелетная) мышечная ткань образует мышцы, прикрепляющиеся к костям скелета. При сокращении (укорочении) скелетных мышц, функции которых подчиняются осознанным усилиям воли человека, кости (костные рычаги) выполняют заданные движения. Исчерченная (скелетная) мышечная ткань образована мышечными волокнами, которые в отдельных мышцах могут достигать в длину 10—12 см. Снаружи каждое мышечное волокно покрыто оболочкой — сарколеммой, в которую вплетаются тонкие коллагеновые волокна, получившие название эндомизия. Под сарколеммой в каждом мышечном волокне, в его цитоплазме (саркоплазме), располагаются многочисленные ядра (до 100), специальные органеллы (миофибриллы), а также органеллы общего назначения и включения (миоглобин, гликоген). Миоглобин, растворенный в саркоплазме, является пигментосодержащим белком, близким по своим свойствам гемоглобину эритроцитов, придающим мышцам красный цвет.
Основную часть мышечного волокна составляют специальные органеллы — миофибриллы (рис. 12). Миофибриллы образованы нитями сократительных белков миозина и актина, расположенными ндоль мышечного волокна в определенном порядке. Эти белковые нити (миофиламенты) скреплены при помощи особых периодически повторяющихся структур, получивших название телофрагма и мезофрагма. Телофрагмы образованы белковыми молекулами, ориентированными поперек мышечного волокна и прикрепленными к сарколемме (оболочке волокна). На продольном срезе мышечного волокна телофрагмы имеют вид темных поперечных линий толщиной около 100 нм, получивших название Z-линий. На середине между двумя соседними телофрагмами располагается также поперечная структура — мезофрагма, на продольном срезе волокна ее называют М-линией.
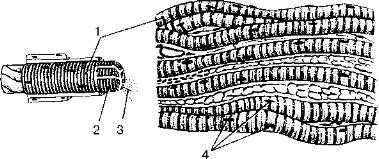
Рис. 12. Исчерченная (поперечнополосатая, скелетная) мышечная ткань:
1 — мышечное волокно; 2 — сарколемма; 3 — миофибриллы; 4 — ядра
От мезофрагмы в сторону телофрагмы отходят тонкие (5 нм) актиновые нити. Навстречу этим нитям от телофрагмы идут толстые (10 нм) миозиновые нити, проникающие между актиновы- ми нитями.
Участок между двумя Z-линиями (телофрагмами) называют саркомером, который является структурно-функциональной единицей миофибриллы. Часть миофибриллы, занятая мезофрагмой (М-линией) с отходящими от него в обе стороны миозиновыми нитями (миофиламентами), получила название Н-полосы (светлая зона). Та часть миофибриллы, в которой располагаются и нити миозина, и нити актина, является А-полоской (A-диск). Части двух соединенных саркомеров, занятые Z-линией (телофрагмой)
с отходящими от нее в обе стороны актиновыми нитями, образуют j-полоску (j-диск).
Чередование темных A-дисков и светлых j-дисков, располагающихся на одном уровне в соседних миофибриллах, создает на гистологическом препарате скелетной мышцы впечатление поперечной исчерченности. Сарколемма на уровне телофрагмы образует глубокие впячивания, в которых располагаются поперечные трубочки (Т-трубочки) незернистой эндоплазматической сети, разветвляющиеся между миофибриллами мышечного волокна.
В основе мышечного сокращения лежат взаимодействия между актином и миозином. При сокращении мышцы актиновые мио- филаменты скользят навстречу миозиновым миофиламентам. При расслаблении мышцы миофиламенты двигаются в противоположные стороны. При этом длина дисков А не изменяется, а диск] — уменьшается в размерах.
По количеству миофибрилл в саркоплазме мышечные волокна подразделяются на медленные («красные»), содержащие мало миофибрилл и много саркоплазмы, и быстрые («белые»), в которых много миофибрилл и мало саркоплазмы. «Красные» мышечные волокна медленно сокращаются, но могут быть долго в рабочем состоянии. «Белые» мышечные волокна быстро сокращаются и быстро устают. Сочетание в мышцах медленных и быстрых исчерченных (поперечнополосатых) мышечных волокон обеспечивает их быстроту реакции (сокращения) и длительную работоспособность.
Источником развития поперечнополосатой (скелетной) мышечной ткани являются клетки миотомов сомитов. На ранних стадиях развития зародыша из мезодермы миотомов выселяются одноядерные веретенообразные клетки — миобласты. Быстро размножаясь, миобласты в соответствующих местах образуют закладки будущих мышц. Быстрое деление ядер приводит к утрате миобластами клеточного строения и они превращаются в крупные многоядерные комплексы — мышечные волокна. В формирующихся мышечных волокнах увеличивается количество миофибрилл, появляется поперечная исчерченность. Во второй половине внутриутробного развития и постнатальном онтогенезе мышечные волокна растут в длину и толщину путем увеличения числа содержащихся в них миофибрилл. Вместе с ростом и дифферен- цировкой мышечных волокон происходит слияние их с клетками-сателлитами. Клетки-сателлиты располагаются под сарколеммой мышечных волокон и являются источником новых волокон. Клетки-сателлиты способны делиться и давать начало миоблас- там после мышечной травмы.
Неисчерченная (гладкая) мышечная ткань образует сократимый аппарат в стенках внутренних органов, протоков желез, кровеносных и лимфатических сосудов и других органов. Структурным gt;лементом этой ткани Являются гладкомышечные клетки (миоци- ты). Гладкие миоциты представляют собой, веретенообразной формы клетки длиной 20—500 мкм, толщиной 5—8 мкм. Каждый ми- оцит имеет одно палочковидное ядро, расположенное в середине клетки. При сокращении миоцита ядро изгибается и даже спира- I с видно закручивается. Органеллы, в том числе и многочисленные митохондрии, расположены ближе к полюсам клетки. Эндоплазматическая сеть и комплекс Гольджи развиты слабо, что свидетельствует о низкой синтетической функции миоцитов. I? цитоплазме миоцитов много актиновых и миозиновых фибрилл, расположенных не параллельно, а под углом одна к другой. Доля актина (по сравнению с миозином) в гладких миоцитах выше, чем в исчерченных (поперечнополосатых) мышечных волокнах. Взаимодействие актиновых и миозиновых миофибрилл происходит по принципу скольжения, но осуществляется оно иначе, чем Igt; скелетной мышечной ткани. Гладкие миоциты не имеют поперечнополосатой исчерченности, сокращаются они помимо усилия ноли, их функции находятся под контролем автономной (вегета- I ивной) части нервной системы. Гладкие миоциты обьединяются в пучки, в образовании которых участвуют тонкие коллагеновые и (ластические волокна.
Сердечная исчерченная мышечная ткань образована плотно прибегающими одна к другой, имеющими поперечнополосатую испорченность мышечными клетками — кардиомиоцитами. В то же нремя сердечные мышечные клетки сокращаются автоматически, подчиняясь ритму проводящей системы сердца и функциям автономной (вегетативной) нервной системы. Кардиомиоциты представляют собой удлиненные (до 100—150 мкм) клетки толщиной 10—20 мкм, каждая из этих клеток имеет ядро, расположенное в ; центре. Органеллы общего значения располагаются в области конца клетки. Митохондрии располагаются цепочками вдоль миофибрилл. В кардиомиоцигах имеются включения — гликоген, липиды. !? кардиомиоцитах актиновые и миозиновые миофибриллы распо- !агаются так же, как в клетках скелетной мускулатуры. Тонкие актиновые миофибриллы одним концом прикреплены к тело- фрагме, образующей линию Z. Толстые (миозиновые) миофибриллы, расположенные между актиновыми, одним своим кондом прикрепляются к мезофрагме (линии М), а другим направлены г. сторону телофрагмы.
Кардиомиоциты, контактируя один с другим, образуют в струк- |урном и функциональном отношениях целостную сократитель
ную систему. На границе прилегающих один к другому кардиоми- оцитов находятся вставочные диски, состоящие из соприкасающихся участков цитолеммы контактирующих клеток, наподобие расширенных десмосом. Вставочные диски прочно соединяют соседние кардиомиоциты и в то же время обеспечивают быстрое прохождение через них нервных импульсов, что дает возможность всем сердечным миоцитам сокращаться одновременно. С помощью вставочных дисков обеспечивается не только структурное, но и функциональное объединение кардиомиоцитов в целостную сердечную мышцу (миокард).