
Анеуплоидия — (трисомия и моносомия) — возникает в результате нарушения сегрегации хромосом в митозе или в мейозе.
Классификация механизмов нерасхождения хромосом достаточно условна, так как все процессы, обеспечивающие клеточное деление взаимосвязаны и взаимозависимы.
В широком смысле, термин «нерасхождение» относится к любым нарушениям разделения хроматид или гомологичных хромосом в анафазе митоза или мейоза. Как правило, аномальное расхождение затра
гивает одну пару хромосом. Нерасхождение нескольких пар хромосом, соответственно, приводит к множественным анеуплоидиям.
- основные механизмы возникновения анеуплоидии
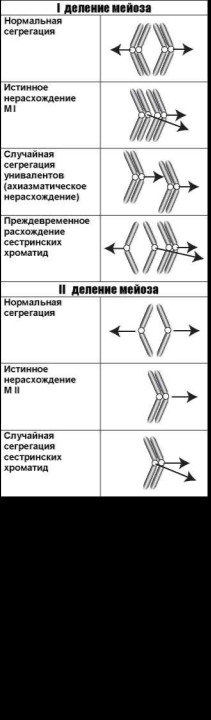
Ниже рассмотрены основные механизмы возникновения анеуплоидии на примере аномальной сегрегации хромосом в мейозе (рис. 3.2).
- собственно нерасхождение хромосом
Собственно нерасхождение хромосом приводит к образованию дочерних анеуплоидных клеток — гипер- и гипоплоидных — вследствие сегрегации обеих гомологичных хромосом (хроматид) к одному полюсу в анафазе клеточного деления (истинное нерасхождение).
- Предделение
Предделение — преждевременное разделение центромер. Предделение мейотических хромосом (бивалентов), как правило, сопровождается образованием унивалентов. Сегрегация унивалентов к разным полюсам регистрируется как нормальная сегрегация, к одному полюсу — как нерасхождение. В качестве одного из механизмов аномальной сегрегации хромосом в оогенезе рассматривается преждевременное разделение сестринских хроматид в первом делении мейоза [424]. Хромосомный набор во втором полярном тельце и ооците в этом случае будет определяться случайной комбинацией одиночных сестринских хроматид.
Преждевременное разделение хроматид и разрыхление центромерных районов хромосом при митотических делениях, сопровождающееся повышенной частотой образования анеуплоидных соматических клеток и микроядер, является цитогенетической характеристикой синдрома Робертса (OMIM 268300). Разъединение сестринских хроматид в центромерном районе одной из Х-хромосом — относительно частое явление, наблюдаемое в отдельных 47,ХХХ лимфоцитах периферической крови у женщин с привычным невынашиванием [300].
- Запаздывание хромосом
Запаздывание хромосом (anaphase lagging) — это полное отсутствие или замедленное относительно остальных хромосом движение хромосомы (или хроматиды) в анафазе, обусловленное, как прави-
ло, нарушением ее ориентации. По-видимому, ведущую роль в этом механизме играют нарушения микротрубочек веретена деления [396], формирования и функций кинетохора — части центромеры, ответственной за прикрепление микротрубочек митотического веретена [763, 793]. Обычно запаздывающие хромосомы элиминируются или образуют микроядра, что сопровождается моносомией при митотических делениях [396] или приводит к гипогаплоидии при мейотических делениях [535].
- Первичное и вторичное нерасхождение хромосом
Нерасхождение хромосом принято подразделять на первичное и вторичное [134].
Под первичным нерасхождением понимают аномальную сегрегацию хромосом в случае вступления в мейоз нормальной диплоидной клетки. Первичное нерасхождение в свою очередь подразделяют на три типа.
- Простое нерасхождение, результатом которого являются гаметы с нуллисомией или дисомией.
- Двойное нерасхождение, к которому относят сочетание (комплементацию) двух простых нерасхождений, одно из которых произошло в оогенезе, другое — в сперматогенезе.
- Последовательное нерасхождение, приводящее к возникновению полисомий, является результатом двух событий нерасхождения одной и той же пары хромосом — сначала в первом, а затем и во втором делениях мейоза.
Нерасхождение хромосом в гаметогенезе у трисомного субъекта называется вторичным нерасхождением.
На многих модельных объектах твердо доказано, что существенную роль в нарушении сегрегации хромосом играет мейотическая рекомбинация, а также хорошо изучены гены, мутации в которых способствуют аномальной сегрегации хромосом. Механизмы возникновения анеуплоидии у человека, в отличие от модельных объектов, менее изучены, а «мейотические» мутации, влияющие на расхождение хромосом у человека, до сих пор не идентифицированы. Тем не менее, накопленные к настоящему времени факты свидетельствуют о сходстве механизмов аномальной сегрегации хромосом в мейозе у различных видов эукариот.
- роль рекомбинации в нерасхождении хромосом
Для исследования связи между рекомбинацией и сегрегацией хромосом у человека используют сравнение частот и локализации обменов и генетических карт сцепления в нормальном мейозе и при нерасхождении хромосом. Именно такой подход позволил доказать, что число и локализация хиазм (обменов) у дрозофилы находятся под генетическим контролем, который запрещает рекомбинацию в прицентромер- ных или теломерных участках, поскольку проксимальное и дистальное расположение хиазм препятствует нормальной сегрегации хромосом.
Наиболее детально изучена взаимосвязь между частотой и локализацией хиазм и нерасхождением хромосом 21 как у матерей, так и у отцов детей с синдромом Дауна [548]. Напомним, что нерасхождение хромосом 21 в сперматогенезе объясняет только 5-10 % случаев синдрома Дауна, а соотношение ошибок сегрегации в первом и втором делениях мейоза близко к 1:1 [210]. Оказалось, что при нерасхождении отцовских хромосом в первом делении мейоза частота хиазм снижается, а во втором делении — соответствует нормальной [401]. В ооге- незе, при трехкратном преобладании ошибок в первом делении мей- оза [210], примерно половина случаев нерасхождения хромосом 21 в первом делении обусловлена полным отсутствием хиазм в биваленте, тогда как в остальных случаях оно сопряжено с наличием обменов в субтеломерных районах. При нерасхождении во втором делении обмены преимущественно регистрируются в прицентромерных участках хромосом 21 [292].
Аналогичные данные о локализации обменов были получены также при анализе 6 других типов нерасхождения (случаи 47,XXY отцовского происхождения, случаи трисомии половых хромосом, а также аутосом 16, 18 и 21 и однородительской дисомии 15 материнского происхождения) [548]. Сходные механизмы нарушения сегрегации хромосом в мейозе, обусловленные их рекомбинацией, предполагаются и для других аутосом [460], в том числе и для акроцентрических хромосом групп D и G [514].
Следовательно, нерасхождение хромосом в первом делении мейо- за характеризуется подавлением рекомбинации и преимущественной локализацией хиазм в дистальных участках. При нерасхождении во втором делении мейоза большая часть рекомбинаций регистрируется в проксимальных районах [401, 548, 675, 823]. Увеличение числа теломерных хиазм с большей вероятностью приведет к нерасхождению в первом делении, а прицентромерных — во втором делении мейоза [549].
Установлено, что ахиазматические биваленты подвержены аномальной сегрегации хромосом в первом делении мейоза, а проксимальные и дистальные обмены малоэффективны в обеспечении правильного расхождения хромосом. Полагают, что биваленты утрачиваются, если их удержание происходит только за счет дистальной хроматид- ной конъюгации [692]. Относительно влияния проксимальных хиазм на поведение бивалентов предложено две модели [292, 458]. Первая предполагает, что гомологичные хромосомы не разделяются в анафазе I, что ведет к утрате бивалента или его сегрегации к одному полюсу. Согласно другой модели, реализация проксимальных хиазм приводит к преждевременному расхождению хроматид и их независимой сегрегации к полюсам в анафазе первого или второго деления мейоза.
В любом случае, нерасхождение хромосом в мейозе не только зависит от рекомбинационных событий, происходящих в профазе I, но и определяется ими [823]. Результаты этих исследований легли в основу «двухударной» гипотезы возникновения трисомии, согласно которой первым «уязвимым» событием является формирование бивалентов и образование хиазм в профазе I и вторым — процесс их разделения в первом или втором делении мейоза [458].
- Генетический контроль мейоза
В настоящее время известен целый ряд генов, экспрессия которых осуществляется только в мейозе [895]. Мутации в этих генах препятствуют созреванию гамет, блокируя клетки на тех или иных стадиях профазы I мейоза [225, 796]. Для некоторых из этих генов, идентифицированных у дрозофилы и дрожжей, установлено существование ортологов, то есть аналогов у других видов, что позволяет высказать предположение об их функциональной гомологии с генами, контролирующими оогенез и сперматогенез у млекопитающих [396]. Следует отметить, что многие из них относятся к семейству циклинов, осуществляющих контроль клеточного цикла в митозе и мейозе [895]. Вместе с тем, низкая частота анеуплоидии в сперматозоидах по сравнению с таковой в ооцитах указывает на различия в механизмах контроля клеточного цикла и процессов презиготической селекции гамет в оогенезе и сперматогенезе [489, 499].
Так, критическими для нормальной сегрегации хромосом в анафазе являются завершение репликации, расположение хромосом на экваторе и прикрепление кинетохор к микротрубочкам веретена. Для большинства митотически делящихся клеток, а также для мейотиче- ских делений сперматоцитов незавершенность одного из этих событий приводит к блоку клеточного цикла на соответствующей стадии и зачастую к массовой гибели всего клона анеуплоидных клеток [395].
Нарушение процессов коньюгации и рекомбинации в сперматогенезе приводит к образованию унивалентов, к остановке мейоза в метафазе первого деления и, в конечном счете, — к гибели клетки. Таким образом, в сперматогенезе функционируют эффективные механизмы селекции аберрантных сперматоцитов, что приводит к элиминации гамет, в том числе и потенциально анеуплоидных [574].
В оогенезе мейоз протекает независимо от наличия унивален- тов или нарушений формирования веретена, что свидетельствует об отсутствии характерной для сперматогенеза селекции гамет с хромосомными аберрациями [226]. В отличие от сперматогенеза блок мейоза на стадии метафазы I является обязательным компонентом оогенеза у большинства животных и у человека (см. главу 1). Утрата в ооге- незе и на первых делениях дробления зиготы механизмов селекции, свойственных делящимся соматическим клеткам и сперматоцитам, по всей видимости, и является причиной высокой частоты анеуплоидии в ооцитах и ранних эмбрионах человека [489, 549] (см главу 5).