
В подавляющем большинстве случаев структурные хромосомные мутации передаются потомству одним из родителей, в кариотипе которого присутствует сбалансированная хромосомная перестройка, то есть являются частью сегрегационного генетического груза. Наличие в кариотипе родителей сбалансированных хромосомных перестроек (транслокации, инверсии) неминуемо сопряжено с нарушениями сегрегации соответствующих хромосом в мейозе, следствием чего является закономерное возникновение генетически несбалансированных гамет и, соответственно, появление зародышей с делециями и дупликациями соответствующих сегментов аберрантных хромосом. Частота возникновения генетически неполноценных гамет зависит от типа хромосомных перестроек, пола носителя и некоторых других факторов, более подробно рассмотренных в главах 4 и 6.
Значительно реже структурные аберрации возникают de novo в гаметогенезе, либо в клетках зародыша (мутационный генетический груз). В последнем случае они могут быть представлены мозаичной формой.
Основным механизмом возникновения структурной перестройки любого типа является наличие разрывов в одной или нескольких хромосомах с последующим воссоединением образовавшихся фрагментов.
Разрывы могут возникать в интерфазном ядре как до, так и после репликации ДНК. В результате разрыва, возникшего до репликации (в периоде G1), поврежденными оказываются обе хроматиды метафазной хромосомы (хромосомный, или изохроматидный, разрыв), в фазе S или после репликации — только одна хроматида (хроматидный разрыв).
Единичный хромосомный разрыв, не затрагивающий центромеры, приводит к появлению делетированной хромосомы и фрагмента,
который, будучи ацентрическим (лишенным центромеры), обычно утрачивается в следующем делении клетки. При наличии двух точек разрыва происходит либо восстановление (репарация) целостности хромосомы, либо образование различных типов перестроек, как внутрихромосомных (если разрывы локализованы в одной хромосоме), так и межхромосомных (при наличии разрывов в разных, гомологичных и негомологичных, хромосомах).
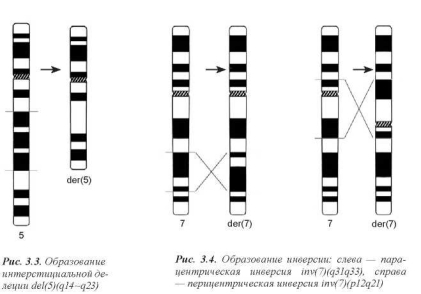
При воссоединении точек разрыва, локализованных в одной хромосоме, может происходить утрата участка хромосомы — делеция (рис. 3.3). Следует отметить, что наличие двух точек разрыва необходимо при образовании интерстициальной делеции, то есть утраты внутреннего сегмента. Для образования концевой, или терминальной, делеции (утраты дистального участка хромосомы) достаточно одного разрыва.
Воссоединение двух точек разрыва в той же хромосоме может происходить с разворотом «вырезанного» участка на 180° (инверсии) (рис. 3.4).
Для образования инсерции (встраивание между точками разрыва участка другой хромосомы) необходимо наличие трех разрывов — один разрыв на хромосоме-«реципиенте» и два других — для образования участка-«донора». Инсерция может осложняться инверсией «донорского» участка (рис. 3.5).
При образовании дупликации воссоединение точек разрыва сопровождается удвоением какого-либо участка хромосомы. Если удвоенные участки располагаются последовательно, то такая дупликация называется тандемной, если с разворотом на 180° — инвертированной (рис. 3.6).
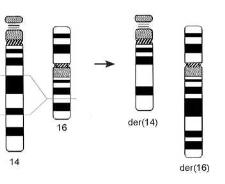
Рис. 3.5. Образование инсерции der(16) вследствие межхромосомной перестройки. Хромосома-«реципиент» — прямая инсерция dir ins(16;14)(q22;q21q23). Второй продукт перестройки (хромосома-«донор») — del(14)(q21q23)
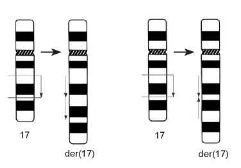
Рис.3.6. Образование дупликации. Слева — тандемная дупликация dup(17)(q21q23), справа — инвертированная дупликация dup(17)(q23q21)
Если утрачиваются оба теломерных участка одной хромосомы, то воссоединение открытых концов сопровождается образованием кольца. Судьба кольцевой хромосомы может быть различна (рис. 3.7).
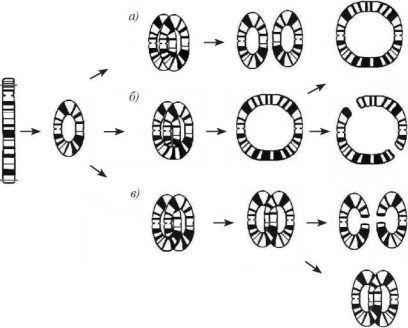
Рис. 3.7. Образование кольцевой хромосомы и сценарии ее судьбы в зависимости от особенностей репликации [187]. При нормальной репликации (a) образуется 2 моноцентрических кольца (r); при одном обмене (или нечетном числе обменов) между сестринскими хроматидами (б) — большое дицентрическое (dicr) кольцо; при двух обменах (или их четном числе) (в) — 2 «сцепленных»моноцентрических кольца. Отдельные моноцентрические хромосомы сегрегируют в дочерние клетки без затруднений. Дицентрические и сцепленные моноцентрические кольца в анафазе обычно разрушаются и либо утрачиваются, либо видоизменяются в зависимости от вариантов воссоединения новых точек разрыва и репликации в следующем клеточном цикле
Если разрывы затрагивают разные хромосомы, то обмен фрагментами возможен двояким образом (рис. 3.8, 3.9). В случае сохранения центромеры в каждой из перестроенных хромосом, их сегрегация в митозе происходит без каких-либо затруднений (рис. 3.8). Однако возможно образование двух ацентрических фрагментов и дицентрической хромосомы, судьба которой определяется особенностями поведения в анафазе (рис. 3.9).
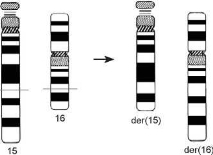
Рис. 3.8. Образование реципрокной транслокации t(15;16)(q22;q22)(15pter^15q22::16q22- 16qter; 16pter^16q22::15q22^15qter)
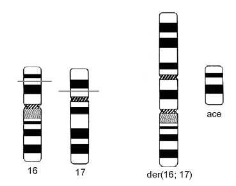
Рис. 3.9. Образование дицентрической хромосомы dic(16;17)(p13.1;p11.2) и ацентрического фрагмента
Для объяснения возникновения изохромосом можно предположить два механизма (рис. 3.10).
- Первый механизм — удвоение плеча одной хромосомы (длинного или короткого) вследствие аномального (поперечного) разделения центромеры.
- Второй механизм предполагает образование изохроматидного разрыва в прицентромерном районе, в результате которого возникают два центрических фрагмента, с последующим восстановлением двуплечей хромосомы из одного плеча и утратой второго плеча.
В определенном смысле изохромосома представляет собой гигантскую инвертированную дупликацию размером с целое плечо и делецию другого плеча. Возникающие вследствие этих внутрихромосомных перестроек изохромосомы являются истинными, так как оба плеча генетически идентичны. Именно этот способ образования изохромосомы считается наиболее распространенным.
В случае межхромосомной перестройки, то есть обмена между идентичными плечами гомологичных хромосом в центромерном районе (или разрыва в центромерах двух гомологичных хромосом с воссоединением двух одинаковых плеч по типу центрического слияния) возникает хромосома, которая морфологически не отличается от изохромосомы, однако ее плечи не являются генетически идентичными (рис. 3.11).
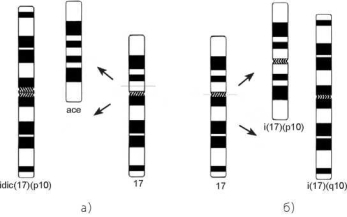
Рис. 3.10. Образование изохромосомы путем изохроматидного разрыва (а) или поперечного деления центромеры (б)
Большинство изохромосом оказываются дицентрическими, однако функции кинетохора выполняет лишь одна центромера, тогда как вторая оказывается репрессированной.
Образование изохромосом может быть результатом митотической рекомбинации, но чаще происходит в гаметогенезе и затрагивает Х-хромо- сому. Если аберрантная Х-хромосома имеет отцовское происхождение, то единственным механизмом ее возникновения может быть слияние генетически идентичных сестринских хроматид. Если изохромосома Х имеет материнское происхождение, то в ее формировании могут принимать участие как одна, так и две Х-хромосомы. В первом случае механизм образования изохромосомы сходен с таковым в сперматогенезе, во втором случае нельзя исключить транслокацию или хрома- тидный обмен между гомологичными хромосомами.
Сложности в определении механизма образования аберрации представляют и аутосомные изохромосомы. Установить природу аберрации (изохромосома или межхромосомный обмен) принципиально возможно молекулярными методами с использованием высокополиморфных маркеров. Истинная изохромосома будет гомозиготна по всем локусам обоих плеч, тогда как гетерозиготность будет свидетельствовать о вовлечении двух хромосом в образование изохромосомы.
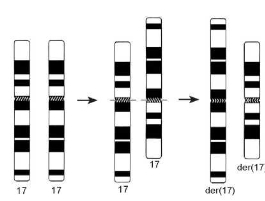
Рис. 3.11. Обмен одноименными плечами гомологичных хромосом в районе центромеры. Образующиеся продукты транслокации морфологически не отличаются от изохромосомы
Разрывы в центромерных районах, сопровождающиеся обменами между целыми плечами разных хромосом или приводящие к разъединению двуплечей хромосомы на две акроцентрические — достаточно редкие события. Значительно чаще обмены в районе центромеры затрагивают акроцентрические хромосомы и приводят к их центрическому слиянию — Робертсоновской транслокации, названной так по имени автора У Робертсона, впервые описавшего этот вид хромосомных перестроек у некоторых видов прямокрылых в 1916 году. Возникшая вследствие слияния двух акроцентриков метацентрическая или субметацентрическая хромосома обычно имеет одну центромеру, реже — две. В последнем случае одна из центромер может быть неактивна (рис. 3.12).
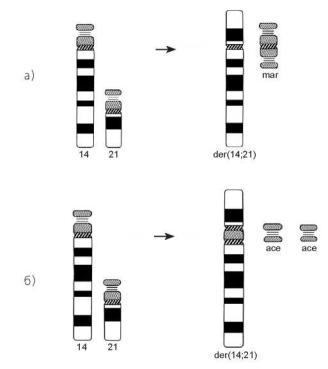
Рис. 3.12. Центрическое слияние (образование Робертсоновской транслокации) с сохранением одной (а) или двух (б) центромер
Короткие плечи акроцентриков могут сливаться с образованием моно- или дицентрической маркерной хромосомы. Иногда утрачивается лишь одно короткое плечо, а второе представлено отдельной мелкой маркерной хромосомой. Оба коротких плеча в виде маркерной хромосомы сохраняются реже. Чаще короткие плечи обоих акроцентриков теряются, поэтому число хромосом у гетерозиготного носителя Робертсоновской транслокации становится на единицу меньше. При этом, однако, учитывая положение точек разрывов в околоцентромерных районах обеих хромосом, реальной утраты генетического материала не происходит.
В центрических слияниях равновероятно могут участвовать все пять пар акроцентрических аутосом. Между тем, отдельные хромосомы групп D и G вовлечены в образование Робертсоновских транслокаций не случайно. Это подтверждается данными о частоте носителей транслокаций D/D, D/G, G/G среди новорожденных (табл. 3.1). Наиболее часто в образовании Робертсоновских транслокаций принимают участие хромосомы 14, 13 и 21.
Таблица 3.1. Частота гетерозиготных носителей центрических слияний разных акроцентрических хромосом [187]
|
Хромосомы |
13 |
14 |
15 |
21 |
22 |
Всего |
|
13 |
|
101 |
8 |
10 |
6 |
125 |
|
14 |
|
|
8 |
45 |
3 |
56 |
|
15 |
|
|
|
17 |
3 |
20 |
|
21 |
|
|
|
|
17 |
17 |
|
22 |
|
|
|
|
|
|
|
Всего |
|
101 |
16 |
72 |
29 |
218 |
Как уже отмечалось, дицентрические хромосомы и кольца с двумя функциональными центромерами обычно нарушают деления клетки. Подавляющее большинство хроматидных разрывов и обменов, возникающих после репликации ДНК, также элиминируются в следующем митозе. Некоторые аберрантные хромосомы принимают участие в образовании специфических структур — микроядер. Микроядерный тест, а также учет мостов и фрагментов в анафазе и хромосомных и хроматидных разрывов и фрагментов в метафазе широко используется при оценке влияния мутагенных факторов на соматические клетки.
Более стабильными являются внутри- и межхромосомные перестройки с сохранением функций одной центромеры. Именно такие хромосомные аберрации совместимы с развитием и живорождением и сравнительно часто регистрируются при пренатальном и постнатальном кариотипировании.
Следует отметить, что точки разрыва могут быть локализованы практически в любом участке любой хромосомы, однако, частота их возникновения в разных сегментах варьирует. Кроме того, типы перестроек для отдельных хромосом проявляют определенную специфику.
Так, для акроцентрических хромосом наиболее характерны центрические слияния (см. выше).
Высокоспецифичной реципрокной транслокацией является Филадельфийская хромосома (аббревиатура Ph или Ph1) — продукт транслокации хромосом 9 и 22 — der(22)t(9;22)(q34;q11), которая закономерно возникает при хроническом миелолейкозе.
Среди аберрантных Х-хромосом лидируют изохромосомы по длинному плечу и кольцевые Х-хромосомы, которые клинически проявляются синдромом Шерешевского-Тернера.
Из аномалий хромосомы 18 наиболее часто встречаются изохромосомы по короткому плечу и кольцевые хромосомы. Каждая из перестроек сопровождается очерченной клинической картиной. Это позволило выделить их в отдельные синдромы, получившие название по типу хромосомной аберрации — синдром i(18p) и синдром r(18).
Подавляющее большинство других типов перестроек (делеций, дупликаций, инверсий, инсерций и реципрокных транслокаций) характеризуются уникальностью точек разрыва и повторяемостью только в пределах одной родословной. В качестве исключения из этого правила можно привести относительно частые спорадические случаи делеций и дупликаций, которые по характерной симптоматике выделены в отдельные синдромы. Так, известны делеционные синдромы:
- короткого плеча хромосомы 4 (синдром Вольфа-Хиршхорна);
- короткого плеча хромосомы 5 (синдром «крика кошки»);
- короткого плеча хромосомы 9 (синдром Альфи), иногда в форме изохромосомы 9 по длинному плечу;
- длинного плеча хромосомы 13 (синдром Орбели);
- короткого и длинного плеч хромосомы 18 (синдромы 18p- и 18q- соответственно);
- длинного плеча хромосомы 21 (синдром 21q-);
- длинного плеча хромосомы 22 (синдром 22q-).
Делеции 13q-, 21q- и 22q- часто приводят к образованию кольцевых хромосом, поэтому их иногда обозначают синдромами кольцевых хромосом 13, 21 и 22.
К разряду «синдромальных» относятся и дупликации сегментов некоторых хромосом:
- короткого плеча хромосомы 9 (синдром 9р+);
- длинного плеча хромосомы 14 (синдром 14q+).
Следует отметить также нозологические формы (всего около 20), обусловленные микроперестройками (делециями и дупликациями). Наиболее известные из них:
- синдромы Прадера-Вилли или Ангельмана в зависимости от родительского происхождения микроделеции в импринтиро- ванном районе 15q11-q13;
- синдром Видемана-Беквита (дупликация сегмента 11р15);
- синдром Лангера-Гидеона (микроделеция 8q24.11-q24.13);
- ретинобластома (микроделеция 13q14);
- Миллера-Диккера (микроделеция 17р13.3);
- синдром Вильямса (микроделеции 7q11 и реже — 11q13-q14; 22q-);
- синдром ДиДжорджи (микроделеция 22q11).
Заключение
В главе рассмотрены основные типы хромосомных перестроек, механизмы их возникновения на уровне всего генома, отдельных хромосом и их сегментов, затронута проблема презиготической селекции гамет. Отмечено, что хромосомные перестройки представляют собой обширный и гетерогенный класс мутаций.
Численные нарушения на уровне генома и целых хромосом возникают обычно в митозе и в мейозе, а также могут быть результатом нерасхождения хромосом на начальных стадиях дробления оплодотворенной яйцеклетки. Механизмы возникновения структурных хромосомных перестроек более многообразны и полиморфны. Обычно первым этапом их возникновения являются хроматидные разрывы, индуцированные мутагенами. Если эти повреждения ДНК не репа- рируются при следующем раунде репликации, возникают хромосомные разрывы, которые посредством рекомбинационного механизма приводят к образованию внутри- или межхромосомных перестроек. При этом ошибки рекомбинации более характерны для мейоза, в котором обмены участками ДНК между бивалентами являются обязательными компонентами нормального процесса созревания гамет, в то время как митотический кроссинговер происходит существенно реже. Частота и спектр хромосомных мутаций в значительной степени определяются спецификой затронутых перестройкой сегментов, в частности, локус- и хромосом-специфичностью, а также другими факторами. Подробнее эти аспекты будут рассмотрены в главах 4, 5 и 6.