5.2. Физиология синапса
В конце XIX в. Ч. Шеррингтон назвал связь между двумя нейронами синапсом. Он же высказал мнение об их функциональной роли, указав, что одни могут являться возбуждающими, а другие — тормозящими.
В настоящее время выделяют две основные разновидности синапсов — химические и электрические. В основу этой классификации положен различный механизм передачи информации в виде нервного импульса от одного нейрона к другим [13,26, 32]. Существуют и смешанные — электрохимические синапсы.
Синапсы классифицируются также в зависимости от того, какими элементами нейрона они образованы (рис. 5.3).
| * | ±z | ~1 |
| Аксо- I | Аксо- | Дендро- [ |
| аксональные ] | соматические | денритические | |
I Аксо-
I дендритические
Классификация синапсов по месту их локализации
Л
Первыми синапсами, которые удалось идентифицировать с помощью электронной микроскопии, были простые контакты терминалей, относимые к аксо-соматическому и аксо-дендритному типам. Несколько позднее были идентифицированы аксо-аксонные и дендро-денд- ритные типы синапсов.
Электрические синапсы более примитивны, и в мозге высших млекопитающий их мало. Морфологически в электрических синапсах пре- и постсинаптическая мембраны тесно прилегают друг к другу. При этом образуется плотный контакт с низким сопротивлением, через который электрический ток проходит из одной нервной клетки в другую. Поскольку сигнал, проходя через синапс, мало модифицируется, то электрический сигнал пресинаптической клетки вызывает сходный стлал в постсинаптической клетке. Таким образом, в электрическом синапсе информация передается без каких-либо химических веществ (медиаторов), а только за счет физических особенностей контакта. Как следствие этого, задержка при проведении возбуждения в электрических синапсах значительно меньше, чем в химических. Однако целый ряд физиологических механизмов, происходящих в химических синапсах, невозможно реализовать в электрическом синапсе. Так, например, передача информации электрическими синапсами не сопровождается следовыми процессами. К тому же такой синапс очень жестко детерминирован и, следовательно, обладает низкой пластичностью. Основным его преимуществом перед химическим является скорость передачи.
Остановимся более подробно на физиологии химического синапса (рис 5.4).
В химическом синапсе выделяют пресинаптическое окончание, синаптическую щель и постсинаптическую мембрану [13,14,23].
Пресинаптическое окончание образуется, как правило, по ходу раз- ветвления аксона, иннервирующего дру1ую клетку. Главным ультра- структурным элементом пресинаптического окончания, обеспечивающим синапическую передачу, являются синаптические пузырьки (везикулы). В пресинаптическом окончании содержится несколько ты сяч везикул, представляющих собой определенную форму упаковки химического вещества, участвующего в передаче влияния через синапс и называемого медиатором. Медиатор образуется либо в теле нейрона и по механизму аксонального транспорта попадает в пресинаптическое окончание, либо синтезируется или ресинтезируется непосредственно на месте. Для синтеза медиатора нужны ферментативные системы, образующиеся в теле клетки на рибосомах и доставляющиеся в пресинаптическое окончание аксональным транспортом.
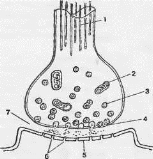
Рио. 5.4. Строение синапса
1 — микротрубочки; 2—митохондрии; 3 — синаптические пузырьки с медиатором; 4 — пресинаптическая мембрана; 5 — постсинаптическая мембрана; 6 — рецепторы; 7 — синаптическая щель.
Синаптическая щель имеет ширину 20-50 нм. В ней содержатся межклеточная жидкость и вещество мукополисахаридной природы в виде тяжей между пре- и постсинаптической мембранами. В синаптической щели также находятся ферменты, некоторые из которых могут разрушать медиатор.
Постсинаптическая мембрана представляет собой часть клеточной мембраны иннервируемой клетки. На мембране расположены белковые рецепторы, чувствительные к медиатору. Рецепторы могут иметь ионные каналы или быть связанными с белками, формирующими такие каналы. Взаимодействие медиатора с белком рецептором приводит к изменению активности ионных каналов.
В процессе передачи сигнала в химическом синапсе можно выделить следующие этапы.
- Потеитциал действия поступает в пресинаптическое окончание.
- После поступления ПД к пресинаптическому окончанию происходит деполяризация мембраны этого окончания, активируются потенциал-зависимые кальциевые каналы и в синаптическую тер- миналь входит Са2+.
- Повышение концентрации ионов Са2+ активирует транспортную систему везикул, что инипиирует их экзоцитоз.
- Содержимое везикул выделяется в синаптическую щель.
- Молекулы медиатора, диффундируя в синаптической щели, связываются с рецепторами постси*1аПтической мембраны.
- Рецепторы постсинаптической мембраны активируют ионные каналы.
- В результате под действием медиатора происходит активация ионных каналов и переход по этим каналам ионов ц0 их градиентам концентрации. Движение ионов формирует Постсинаптиче- ский потенциал, который по своим свойствам является локальным ответом.
- Медиатор, находящийся в контакте с рецепторами постсинаптической мембраны и в синаптической щели, разруЩается фермен. тами.
- Продукты разрушения медиатора и, возможно, не разрушенный медиатор всасываются преимущественно в пресинаптическое окончание, где осуществляется ресинтез медиатора и помещение его в везикулы (в случае если медиаТОр не доставляется аксональным транспортом в пресинаптическое окончание)
На все эти процессы требуется определенное время, которое получило название синаптической задержки и составляет 0 2-0 5 мс Синаптическая задержка пропорционально зависит от температуры При активации постсинаптических каналов медиаторами может возникать достаточно сильный ионный ток, способный деполяризовать крупные клетки. Поэтому при химической передаче мелкие Нресинаптические волокна могут возбуждать большие постсинаптическце клетки. Следовательно, химический компонент синаптической Передачи является усилителем электрических сигналов.
Химическая передача осуществляет как возбуждающее, так и тормозное действие на постсинаптическую клетку и зависит от медиатора и рецепторов на постсинаптической мембране. Из пресинаптиче- ского окончания могут выделяться це только медиаторы, но и другие вещества. В связи с этим можно выделить определенные критерии медиатора;
- он должен синтезироваться в нейроне;
- накапливаться в окончании клетки;
- оказывать специфическое действие на постсинаптическую мембрану;
- выделяться при появлении кальциевого тока в пресинаптическое
ПКП1ГМ1П1С*.
В одном синаптическом пузырьке находится порция медиатора, которая называется квант. Помимо выделений медиаторов, инициируемых нервным импульсом, в химических синапсах имеются спонтанные, обычно редкие миниатюрные изменения трансмембранного потенциала в области постсинаптической мембраны, составляющие доли милливольта. Они отражают спонтанный выброс одиночных квантов медиатора и реакцию на них постсинаптической мембраны.
В 30-х гг. XX в. стал известен так называемый принцип Дейла, названный по имени открывшего его ученого — Ч. Дейла. Суть его в следующем: клетка может синтезировать своими терминалями только один медиатор [26]. Позже этот принцип был уточнен: из одного окончания клетки может выделяться не один, а несколько медиаторов, причем этот комплекс постоянен для каждой конкретной клетки. Одно вещество из этого комплекса выполняет функцию медиатора, а остальные могут выполнять функцию модуляторов, т. е. веществ, усиливающих или ослабляющих чувствительность к медиаторам рецепторов постсинаптической мембраны. Некоторые вещества могут выполнять не только функции медиаторов, но и ряд других. К таким веществам могут относиться, например, тахикинины. Группу тахикининов формируют пептиды, имеющие сходную с веществом Р последовательность аминокислот. Сюда относятся соединения, обозначенные как нейро- кинины, бомбезины. Распределение тахикининов в тканях одного и того же организма обеспечивает широкий спектр пх физиологической активности:
- сокращение гладкой мускулатуры,
- формирование поведенических и гормональных процессов,
- высвобождение других физиологически активных веществ,
- периферическую ноцицепцию.
Функциональное разнообразиетахикининов связано с их взаимодействием с другими физиологически активными веществами, как пептидной, так и непептидной природы.
Количество выделяемого медиатора и, следовательно, величина постсинаптического потенциала зависят от амплитуды и частоты потенциала действия, поступающих к окончаниям пресинаптического нейрона [13,27,32].
Таким образом, среди свойств химического синапса выделяют:
- одностороннее проведение возбуждения, которое осуществляется всегда в направлении от пресинаптического окончания в сторону постсинаптической мембраны;
- замедленное проведение сигнала объясняется синаптической задержкой: необходимо время для выделения медиатора из преси- наптического окончания, диффузии его к постсинаптической мембране, возникновения постсинаптического потенциала;
- низкая лабильность синапсов объясняется наличием синаптической задержки и обеспечивает трансформацию ритма возбуждения, поступающего к пресинаптической терминали в отличающийся ритм возбуждения постсинаптической терминали;
- проводимость химических синапсов сильно изменяется под влиянием биологически активных веществ, лекарственных средств и ядов, гипоксии.
В головном мозге относительно редко встречаются изолированные одиночные синапсы. Обычно несколько синапсов, расположенных на определенном участке мембраны, формируют определенный тип групповой синаптической связи. Возможны весьма разнообразные варианты Например, два или несколько синапсов могут быть расположены рядом друг с другом и ориентированы в одном направлении. Их отростки могут образовывать более сложные структуры на поперечном срезе, напоминающие стопку монет. Такие синапсы называют последовательными. Примером могут служить аксо-аксо-дендритные и аксо-дендро-денд- ритные последовательности. Возможен вариант, называемый реципрок- ным синапсом. В этом случае один отросток соединяется с другим отростком, а последний образует синапсы на первом.
Если два таких синапса расположены рядом, то их называют реципрок- ной парой.
Если же два синапса удалены один от другого, то возникает реципрокное устройство.
Есть такие синаптические соединения, когда тесно сближена целая группа терминалей. Этот тип называют синаптической гломерулой.
‘ Таким образом, в зависимости от задач, решаемых при синаптической передаче, формируются различные типы контактов.
Эффективность передачи в синапсе зависит от интервала следования сигналов через синапс [26].
Если учащать подачу импульса по аксону, то на каждый последующий потенциал действия ответ постсинаптической мембраны, выраженный величиной изменения трансмембраниого потенциала в ответ на воздействие, будет возрастать. Это явление облегчает передачу сигнала в синапсе, усиливая ответ постсинаптического нейрона на очередной раздражитель. Подобное явление получило название «облегчение» или «потенциация». В основе этого процесса лежит накопление кальция внутри пресинаптического окончания при достаточно интенсивной стимуляции. Увеличение количества ионов кальция вызывает экзоцитоз большего количества везикул и, следовательно, большего количества медиатора. Таким образом, большее количество рецепторов на постсинаптической мембране будет активировано и откроется больше ионных каналов, что приведет в большему изменению трансмембранного потенциала на постсинаптической мембране.
Если частота возбуждения пресинаптического окончания в течение короткого времени окажется значительной, то в течение 2-5 мин после ее окончания в ответ на одиночный разряд наблюдается рост амплитуды изменения трансмембранного потенциала в постсинаптическом элементе. Механизм этого процесса связывают не только с накоплением кальция в пресинаптическом окончании, по и с фосфорилированием белков. Подобный процесс получил название посттетатчеасая по-, тенциация.
В том случае если изменения сохраняются не несколько минут, а в течение десятков минут или даже дней, то говорят о долговременной по- тещиации. В подобном процессе участвуют сложные метаболические механизмы.
При черезмерном раздражении пресинаптического окончания в нем происходит истощение медиатора, что приводит, согласно описанным выше механизмам, к уменьшению амплитуды трансмембранного потенциала на постсинаптической мембране. Этот процесс называется кратковременная депрессия. Он развивается и взаимодействует с по- сттетанической потенциацией.
В том случае если стимуляция синапса низкочастотна (порядка 1- 5 Гц), то возникает десинхронизация по времени активации пре- и постсинаптической мембраны, что приводит в свою очередь к долговременной депрессии данного синапса.
Различная интенсивность использования синапса приводит к его модификации, вызывая улучшение или ухудшение передачи через него сигнала (рис. 5.5). Синаптическая пластичность имеет важное значение в процессах обучения, памяти, условных рефлексах, забывания.
На посгсинагггичесокой мембране под действием медиатора может происходить два основных процесса, связанных с возбуждением и торможением. Электрофизиологическим субстратом этих процессов являются изменения трансмембранного потенциала постсииаптической мембраны, получившие названия возбуждающего постсинаптического потенциала (ВПСП) и тормозного постсинаптгшческого потенциала (ТПСП).
| Модификации синапса |
Рис. 5.5. Основные модификации синаптической передачи
Возбуждающий постсинаптический потенциал (ВПСП). В синапсах, в которых осуществляется возбуждение постсинаптической структуры, обычно происходит повышение проницаемости для ионов натрия. По градиенту концентрации ионы натрия входят в клетку, что вызывает деполяризацию постсинаптической мембраны. Эта деполяризация получила определенное название, возбуждающий постсинаптический потенциал (ВПСП), который относится к локальным ответам и, следовательно, обладает способностью к суммации. Выделяют временную и пространственную суммацию.
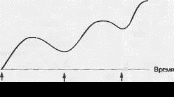
Принцип временной суммации заключается в том, что импульсы поступают к пресинаптическому окончанию с периодом меньшим, чем период ВПСП. Как следствие, новые noprani медиатора выделяются в тот момент, когда трансмембранный потенциал еще не вернулся к уровню МПП. Далее новая деполяризация развивается не с уровня МПП, а с текущего уровня трансмембранного потенциала, который ближе к КУД (рис. 5.6).
КУД
Сущность пространственной суммации заключается в одновременной стимуляции постсинаптической мембраны синапсами, расположенными близко Друг от дру! а. В этом случае ВПСП каждого синапса суммируются (рис. 5.7).
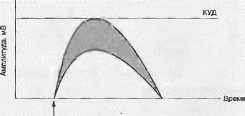
v Рис. 5.7. Пространственная суммация ВПСП
Двойной стрелкой обозначено синхронное возбуждение двух синапсов. Нижний график — ВПСП при возбуждении одиночного синапса, верхний график ВПСП при синхронном возбуждении двух синапсов. Серым цветом выделена область увеличения амплитуды ВПСП.
Если величина ВПСП достаточно велика и достигает критического уровня деполяризации, то генерируется ПД. Однако не все участки мембраны обладают одинаковой способностью кгенерации ВПСП. Так, аксонный холмик, являющийся начальным сегментом аксона относительно сомы, имеет приблизительно в 3 раза более низкий порог электрического раздражения. Следовательно, при равных условиях раздражения именно синапсы, расположенные на аксональном холмике, обладают большей возможностью к генерации ПД, чем синапсы денд- ритов исомы [26,27,32]. От аксонального холмика ПД распространяется в аксон, а также ретроградно в сому. Запись электрогенеза нейрона внутриклеточным электродом позволяет выявить элементы ПД, связанные с аксональным холмиком и сома-дендритным комплексом.
Среди нейронов ЦНС у разных животных (а также среди элементов сетчатки глаза человека) имеются и такие, возбуждение которых исчерпывается постсинаптическим электрогенезом. Эти клетки не могут генерировать потенциал действия, но за счет небольших размеров аксонов ВПСП, возникающих в соме, могут электротонически распространяться до окончания аксона, вызывая секрецию медиатора.
Тормозной постсинаптический потенциал (ТПСП). Задачей пост- синаптического торможения является снижение возбудимости мембраны нейрона. Это достигается применением медиаторов, оказывающих тормозящее действие. Например, гамма-аминомасляной кислоты (ГАМК) или глицина. Первая, взаимодействуя с рецептором, открывает в постсинаптической мембране хлорные каналы. Это приводит к движению иона хлора по электрохимическому градиенту. Как следствие, развивается гиперполяризация, в реализации которой принимают участие и выходящие из клетки ионы калия [13,32]. В результате гиперполяризации увеличивается расстояние до КУДа и, следовательно, уменьшается возбудимость.
В тормозных синапсах так же. как и в возбуждающих, помимо вызванных имеются спонтанные миниатюрные ТПСП.
Пресииалтическое торможение. Этим термином в отличие от рассмотренного выше постсинаптического торможения называют снижение или выключение активности синаптического окончания клетки за счет синаптического же торможения, вызванного оканчивающейся на ней терминалью другого нейрона (рис. 5.8). В этих синапсах торможение определяется действием ГАМК, открывающей хлорные каналы на тормозимой терминали. На определенном участке аксона возникает зона торможения, проходя через которую ПД теряет свою амплитуду [13]. В результате синаптическая передача импульса ослабляется или исключается, поскольку снижение амплитуды ПД приводит к снижению выброса медиатора из пресинаптического окончания При одиночном импульсе тормозного аксона длительность пресинаптического торможения составляет 6-7 мс. Следовательно, максимальный тормозной эффект получается, если тормозной импульс достигает аксон-аксо- нального синапса за несколько миллисекунд до прихода сюда ПД возбуждающего аксона. А сам тормозной синапс находится вблизи возбуждающей терминали.
Пресинаптическое торможение предупреждает (или заранее исключает) развитие постсинаптического возбуждения, но оно не может повлиять на него, когда оно уже развито. При пресинаптическом торможении в постсинагггической клетке не обнаруживается ни ТПСП, ии изменений электровозбудимости.
Механизм пресинаптического торможения представлен в различных отделах ЦНС позвоночных и беспозвоночных.
Источник: Щербатых Ю. В., Туровский Я. А., «Физиология центральной нервной системы для психологов» 2007