5.3. Передача потенциала действия по нервным волокнам
При передаче информации между различными участками нервной системы, отстоящими Друг от друга на значительные расстояния, необходимо распространение нервных импульсов (ПД) по аксонам нейронов. При деполяризации мембраны натриевые потенциал-зависимые каналы открываются и Na+ входит в клетку. Следовательно, ток ионов приводит к возникновению тока в возбужденном участке аксона. Под действием ПД электротонически происходит деполяризация соседних участков аксона, и если она достигает КУД, то развивается потенциал действия. Таким образом, ПД перемещается от возбужденного участка к невозбужденному. Причем, имеет место двухстороннее проведение ПД, который распространяется по аксону как антероградно (от тела нейрона к синапсам аксона), так и ретроградно (от аксона к телу и далее к дендритам). ПД обладает свойством затухания, т. е. чем дальше от участка возбуждения, тем ниже его амплитуда. Следовательно, на определенном удалении деполяризовать мембрану до КУДа невозможно из-за уменьшения амплитуды ПД. Изменение амплитуды ПД при электротоиическом распространении характеризуется постоянной длиной мембраны (расстояние, при котором амплитуда ПД уменьшается на 37% от исходной) [13,26,32]. Следовательно, для распространения нервных импульсов по аксону наиболее важны электрическая возбудимость натриевых потенциал-зависимых каналов мембраны ак-
сота и пассивные кабельные свойства аксона (сопротивления и емкости мембраны, сопротивление цитоплазмы). Сам аксои можно представить в виде электрического кабеля, где сопротивление «сердцевины», т. е. цитоплазмы, гораздо меньше, чем «изоляции»-мембраны.
Очевидно, что чем выше скорость распространения нервных импульсов по аксонам, тем более быстро может наступить реакция на раздражитель, что принципиально важно для процессов адаптации. В процессе филогенеза живые организмы выработали два способа увеличения скорости распространения импульса.
Первый из них заключается в увеличении диаметра аксона. При этом снижается сопротивление нервного волокна (типичным примером могут быть гигантские аксоны кальмаров, на которых и проводились исследования). Гигантские аксоны развились в пропессе эволюции у некоторых видов животных для того, чтоб.ы обеспечивать быструю синхронную активацию двигательных рефлексов.
Второй способ увеличения скорости проведения нервных импульсов, реализовавшийся только у позвоночных животных, состоит в повышении сопротивления мембраны. Роль изолятора для кабеля нервного волокна выполняют миелиновые оболочки, сформированные щвановскими клетками. В ходе онтогенеза миелин откладывается как вокруг периферических, так и центральных аксонов. В результате вокруг волокон образуется плотная многослойная оболочка из клеточных мембран. Однако размер олигодендроцита гораздо меньше длины аксона и. следовательно, есть участки, где миелиновая оболочка отсутствует. Эти участки называются перехваты Ршвъе. Особенность их строения заключается в том, что на этих участках возбудимая мембрана аксона контактирует с внеклеточной средой и концентрация потенци- ад-зависнмых Na^-каналов значительно больше, чем на участках, покрытых миелином, где их практически нет. Отсюда следует, что изолирующие свойства миелиновой оболочки обеспечивают рост постоянной длины аксона. Следовательно, ПД может электротонически вызывать активацию натриевых потенциал-зависимых каналов на большем расстоянии и, следовательно, на большем расстоянии вызвать генерацию нового ПД. При этом его скорость будет соответствовать скорости света в среде. ПД вызывает деполяризацию мембраны в области соседних перехватов Ранвье, следовательно, импульсы в таких аксонах возникают лишь в небольших участках мембраны. Новый процесс деполяризации восстанавливает амплитуду ПД, уменьшившуюся при передаче на расстояние, до прежнего уровня, а возникший иа данном перехвате ПД может электротоиически возбудить мембрану на дру
гих перехватах. Так формируется сальтаторное (скачкообразное) проведение, при котором импульсы распространяются, «перепрыгивая» от перехвата к перехвату (рис. 5.9).
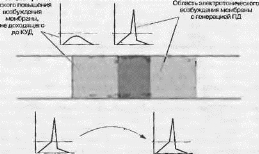
Скорость распространения нервных импульсов зависит от наличия или отсутствия миелиновой оболочки, а также от ее толщины. Скорость распространения импульсов в различных аксонах варьирует от 120 м/с (в толстых миелиновых волокнах) до 50 см/с (в очень немиелинизиро- ванных аксонах). Чем толще миелиновая оболочка, тем выше постоянная длины волокна, и тем дальше могут распространяться ЦД, и тем дальше от места его возникновения происходит деполяризация мембраны до КУДа, и тем выше скорость распространения импульса.
щиеся по электровозбудимости и по скорости проведения (все эти показатели в ряду А-В-С падают):
В свою очередь группа А неоднородна, в ней, по описанным выше показателям, можно выделить подгруппы:
Волокна типа Аа проводят возбуждение от моторных нервных центров спинного мозга к скелетным мышцам (двигательные волокна) и от определенных рецепторов мышц к соответствующим нервным центрам.
Волокна типа Ар, Ау, АА относятся преимущественно к чувствительным, проводящим возбуждение от различных рецепторов (тактильных, температурных, некоторых болевых, рецепторов внутренних органов) в ЦНС. Исключение составляют лишь гамма-волокна, значительная часть которых проводит возбуждение от клеток спинного мозга к инт- рафузальным мышечным волокнам.
К волокнам типа В относятся миелинизированные преганглионар- ные волокна вегетативной нервной системы.
К волокнам типа С относятся безмиелиновые нервные волокна. Это постганглионарные волокна симпатического отдела вегетативной нервной системы, а также нервные волокна, которые проводят возбуждение от болевых рецепторов, некоторых терморецепторов и рецепторов давления.
5.4[1] Аксональный транспорт
Аксоны, помимо проведения возбуждения, являются транспортными каналами для различных веществ. Эти вещества, попав в нейрон или будучи в нем синтезированы, могут перемещаться по аксону как
к его окончанию от тела клетки (антероградный транспорт), так и к телу клетки (ретроградный транспорт) [26, 32]. Перемещаться могут не только отдельные вещества, но и органеллы: митохондрии, везикулы. Существует два вида аксонного транспорта — быстрый и медленный.
Быстрый аксонный транспорт — это транспорт везикул, митохондрий и некоторых белковых частиц со скоростью 250-400 мм/сут. Он осуществляется специальным транспортным механизмом — при помощи микротрубочек и нейрофиламентов и сходен с механизмом мышечного сокращения. Этот транспорт перемещает лизосомы, везикулы, возникающие в окончаниях аксона со скоростью 220 мм/сут, от терминалей аксона к телу клетки. Скорость быстрого антероградного и ретроградного транспорта не зависит от типа и диаметра аксона.
Медленный аксонный транспорт обеспечивает перемещение со скоростью 1—4 мм/сут белков и структур цитоплазмы в дистальном направлении за счет интенсивности синтетических процессов в перикарионе (области цитоплазмы около ядра нейрона). Медленный аксонный транспорт имеет особое значение в процессах роста и регенерации отростков нейрона.
Аксональным транспортом могут пользоваться и патологические агенты: токсины, вирусы, например вирус герпеса.
сота и пассивные кабельные свойства аксона (сопротивления и емкости мембраны, сопротивление цитоплазмы). Сам аксои можно представить в виде электрического кабеля, где сопротивление «сердцевины», т. е. цитоплазмы, гораздо меньше, чем «изоляции»-мембраны.
Очевидно, что чем выше скорость распространения нервных импульсов по аксонам, тем более быстро может наступить реакция на раздражитель, что принципиально важно для процессов адаптации. В процессе филогенеза живые организмы выработали два способа увеличения скорости распространения импульса.
Первый из них заключается в увеличении диаметра аксона. При этом снижается сопротивление нервного волокна (типичным примером могут быть гигантские аксоны кальмаров, на которых и проводились исследования). Гигантские аксоны развились в пропессе эволюции у некоторых видов животных для того, чтоб.ы обеспечивать быструю синхронную активацию двигательных рефлексов.
Второй способ увеличения скорости проведения нервных импульсов, реализовавшийся только у позвоночных животных, состоит в повышении сопротивления мембраны. Роль изолятора для кабеля нервного волокна выполняют миелиновые оболочки, сформированные щвановскими клетками. В ходе онтогенеза миелин откладывается как вокруг периферических, так и центральных аксонов. В результате вокруг волокон образуется плотная многослойная оболочка из клеточных мембран. Однако размер олигодендроцита гораздо меньше длины аксона и. следовательно, есть участки, где миелиновая оболочка отсутствует. Эти участки называются перехваты Ршвъе. Особенность их строения заключается в том, что на этих участках возбудимая мембрана аксона контактирует с внеклеточной средой и концентрация потенци- ад-зависнмых Na^-каналов значительно больше, чем на участках, покрытых миелином, где их практически нет. Отсюда следует, что изолирующие свойства миелиновой оболочки обеспечивают рост постоянной длины аксона. Следовательно, ПД может электротонически вызывать активацию натриевых потенциал-зависимых каналов на большем расстоянии и, следовательно, на большем расстоянии вызвать генерацию нового ПД. При этом его скорость будет соответствовать скорости света в среде. ПД вызывает деполяризацию мембраны в области соседних перехватов Ранвье, следовательно, импульсы в таких аксонах возникают лишь в небольших участках мембраны. Новый процесс деполяризации восстанавливает амплитуду ПД, уменьшившуюся при передаче на расстояние, до прежнего уровня, а возникший иа данном перехвате ПД может электротоиически возбудить мембрану на дру
гих перехватах. Так формируется сальтаторное (скачкообразное) проведение, при котором импульсы распространяются, «перепрыгивая» от перехвата к перехвату (рис. 5.9).
Скорость распространения нервных импульсов зависит от наличия или отсутствия миелиновой оболочки, а также от ее толщины. Скорость распространения импульсов в различных аксонах варьирует от 120 м/с (в толстых миелиновых волокнах) до 50 см/с (в очень немиелинизиро- ванных аксонах). Чем толще миелиновая оболочка, тем выше постоянная длины волокна, и тем дальше могут распространяться ЦД, и тем дальше от места его возникновения происходит деполяризация мембраны до КУДа, и тем выше скорость распространения импульса.
щиеся по электровозбудимости и по скорости проведения (все эти показатели в ряду А-В-С падают):
- группа А включает наиболее толстые, хорошо миелинизирован- ные моторные и чувствительные волокна;
- В слабомиелинизированные преганглинарные (парасимпатические) волокна;
- С — немиелинизированные постганглионарные (симпатические) волокна.
В свою очередь группа А неоднородна, в ней, по описанным выше показателям, можно выделить подгруппы:
- Аа — группа толстых миелиновых волокон, скорость проведения 70-120 м/с;
- Ар — скорость проведения 30-70 м/с;
- Ау — скорость проведения 15—30 м/с;
- АА — скорость проведения 12-30 м/с;
- В — тонкие миелиновые волокна, скорость проведения 5-14 м/с;
- С - безмиелиновые волокна, скорость проведения 0,5-2,3 м/с.
Волокна типа Аа проводят возбуждение от моторных нервных центров спинного мозга к скелетным мышцам (двигательные волокна) и от определенных рецепторов мышц к соответствующим нервным центрам.
Волокна типа Ар, Ау, АА относятся преимущественно к чувствительным, проводящим возбуждение от различных рецепторов (тактильных, температурных, некоторых болевых, рецепторов внутренних органов) в ЦНС. Исключение составляют лишь гамма-волокна, значительная часть которых проводит возбуждение от клеток спинного мозга к инт- рафузальным мышечным волокнам.
К волокнам типа В относятся миелинизированные преганглионар- ные волокна вегетативной нервной системы.
К волокнам типа С относятся безмиелиновые нервные волокна. Это постганглионарные волокна симпатического отдела вегетативной нервной системы, а также нервные волокна, которые проводят возбуждение от болевых рецепторов, некоторых терморецепторов и рецепторов давления.
5.4[1] Аксональный транспорт
Аксоны, помимо проведения возбуждения, являются транспортными каналами для различных веществ. Эти вещества, попав в нейрон или будучи в нем синтезированы, могут перемещаться по аксону как
к его окончанию от тела клетки (антероградный транспорт), так и к телу клетки (ретроградный транспорт) [26, 32]. Перемещаться могут не только отдельные вещества, но и органеллы: митохондрии, везикулы. Существует два вида аксонного транспорта — быстрый и медленный.
Быстрый аксонный транспорт — это транспорт везикул, митохондрий и некоторых белковых частиц со скоростью 250-400 мм/сут. Он осуществляется специальным транспортным механизмом — при помощи микротрубочек и нейрофиламентов и сходен с механизмом мышечного сокращения. Этот транспорт перемещает лизосомы, везикулы, возникающие в окончаниях аксона со скоростью 220 мм/сут, от терминалей аксона к телу клетки. Скорость быстрого антероградного и ретроградного транспорта не зависит от типа и диаметра аксона.
Медленный аксонный транспорт обеспечивает перемещение со скоростью 1—4 мм/сут белков и структур цитоплазмы в дистальном направлении за счет интенсивности синтетических процессов в перикарионе (области цитоплазмы около ядра нейрона). Медленный аксонный транспорт имеет особое значение в процессах роста и регенерации отростков нейрона.
Аксональным транспортом могут пользоваться и патологические агенты: токсины, вирусы, например вирус герпеса.
Источник: Щербатых Ю. В., Туровский Я. А., «Физиология центральной нервной системы для психологов» 2007