
Окончания аксонов периферических нервов подразделяют на чувствительные (афферентные) и двигательные (эфферентные).
А. Чувствительные нервные окончания. Внешние раздражители, а также сигналы о состоянии внутренней среды и двигательной системы организма регистрируют чувствительные нервные окончания — сенсорные рецепторы.
- Классификация. По Лаврентьеву, различают свободные и несвободные воспринимающие приборы.
а. Свободные нервные окончания — терминальные ветвления периферического отростка чувствительного нейрона.
б. Несвободные нервные окончания, помимо терминальных ветвлений периферического отростка чувствительного нейрона, содержат специальные клетки. Практически все окончания этого типа — механорецепторы.
в. Инкапсулированные механорецепторы (инкапсулированные тельца) — несвободные окончания, имеющие оформленную соединительнотканную капсулу.
- Свободные нервные окончания (рис. 8-12) — наиболее распространённый тип сенсорных рецепторов.
а. Локализация. Свободные окончания расположены в прослойках соединительной ткани внутренних органов, а также в соединительнотканной основе кожи, где они сосредоточены преимущественно в сосочках. Свободные нервные окончания эпидермиса расположены в базальном и шиповатом его слоях. В областях кожи с высокой тактильной чувствительностью (например, пальцы рук) терминали достигают зернистого слоя.
б. Строение. Для рассматриваемого вида чувствительных нервных окончаний, локализованных в соединительной ткани, термин свободное условен, т.к. ветвления осевого цилиндра, как правило, окружены вспомогательными клетками (аналогичны шван- новским). Варианты взаимодействия вспомогательных клеток с осевым цилиндром
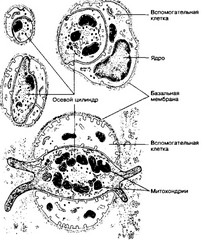
Рис. 8-12. Свободные нервные окончания в соединительной ткани. Осевой цилиндр контактирует с вспомогательными клетками на значительном протяжении или полностью окружён ими. Свободное нервное окончание, изображённое в нижней части рисунка, имеет билатеральную организацию: в центре расположена эллиптической формы нервная терминаль, покрытая вспомогательными клетками [из Andres KH, 1969)
могут быть различны. Так, вспомогательная клетка может покрывать только часть поверхности осевого цилиндра. Остальная его часть отделена от окружающей соединительной ткани лишь базальной мембраной, обычно расположенной по всей поверхности свободного нервного окончания. Нервная терминаль может формировать пальцевидные выросты, проникающие в окружающую соединительную ткань между вспомогательными клетками.
Билатеральная организация. Некоторые свободные окончания в соединительной ткани имеют билатеральную организацию и напоминают сэндвич: в центре расположена нервная терминаль эллиптической формы, а снаружи — вспомогательные клетки. Полагают, что такая организация необходима для регистрации направления механического смещения.
в. Модальность. Большинство свободных нервных окончаний — механорецепторы. Некоторые окончания в эпидермисе определённо специализированы для регистрации изменений температуры. По настоящее время не идентифицированы т.н. холодовые и тепловые окончания. Имеются также рецепторы, способные определять изменения pH, р02 и рС02.
- Комплекс клетки Меркеля с нервной терминалью (рис. 8-13). Осязательные клетки Меркеля — округлые или удлинённые клетки, локализованные в эпителии. Они крупнее эпителиоцитов, ядро вытянутое и дольчатое, цитоплазма светлее, в ней нет характерных для кератиноцитов скоплений промежуточных филаментов. Клетки Меркеля соединяются с эпителиоцитами при помощи десмосом и формируют контакт с нервными терминалями.
а. Структура. В цитоплазме равномерно распределены и в умеренном количестве содержатся митохондрии, лизосомы, мультивезикулярные тельца, вакуоли, микрофиламенты. Эндоплазматическая сеть развита слабо. Встречаются отдельные рибосомы, иногда образующие скопления, гранулы гликогена, меланосомы и центриоли. Между органеллами рассеяно небольшое количество промежуточных филаментов, образующих компактные пучки в пальцевидных выростах. Для клеток Меркеля характерны
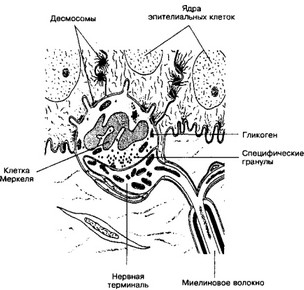
Рис. 8-13. Комплекс клетки Меркеля с нервной терминалью. Клетка Меркеля расположена в базальном слое эпидермиса, образует отростки и связывается с кератиноцитами при помощи десмосом. Расширенная нервная терминаль вступает в контакт с клеткой Меркеля. Специфические гранулы расположены в обращённой к нервной терминали части клетки [из Iggo A, Muir AR, 1969]
специфические осмиофильные гранулы размером от 80 до 200 нм. Они сосредоточены преимущественно в обращённых к нервной терминали участках цитоплазмы. На противоположной от ядра стороне расположен хорошо выраженный комплекс Гольджи, имеющий прямое отношение к формированию специфических гранул.
б. Химия. В клетках Меркеля обнаружены пептиды и нейроноспецифические вещества, что свидетельствует об эндокринной функции клеток Меркеля и позволяет рассматривать их как компонент диффузной нейроэндокринной системы.
- Метионин-энкефалин.
- VIP.
- Относящийся к кальцитониновому гену пептид.
- Вещество Р.
- Пептид гистидин-изолейцин.
- Нейроноспецифическая енолаза.
- Хромогранин А.
в. Нервная терминаль. В области контакта с осязательной клеткой терминаль резко расширена, содержит множество митохондрий, нейрофиламентов, микротрубочек, пузырьков гладкой эндоплазматической сети и липидных включений.
г. Функция. Клетка Меркеля может нести позиционную информацию для определения места окончательной локализации терминали периферического отростка чувствительного нейрона в нейрогенезе или при регенерации нервов.
д. Трансформированные клетки Меркеля присутствуют в нейроэндокринных карциномах кожи и трабекулярных карциномах различной локализации (толстая кишка, лёгкие). В подобных опухолях клетки Меркеля сохраняют некоторые специфичные для них свойства. Так, иммуногистохимическая реакция на нейроноспецифическую енолазу остаётся положительной и служит маркёром диагностики опухолей, содержащих трансформированные клетки Меркеля.
- Тельца Руффйни (рис. 8-14) — крупные рецепторы веретеновидной формы длиной до
-
 мм и диаметром около 150 мкм, расположены в соединительной ткани кожи и суставов.
мм и диаметром около 150 мкм, расположены в соединительной ткани кожи и суставов.
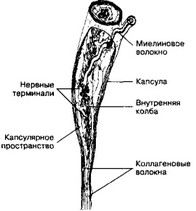
Рис. 8-14. Тельце Руффини содержит внутреннюю колбу с густой сетью разветвлённых нервных терминален и вспомогательными пластинчатыми клетками. Снаружи тельце покрыто соединительнотканной капсулой из нескольких слоев уплощённых фибробластов. Между внутренней колбой и капсулой находится заполненное жидкостью капсулярное пространство [из Chambers MR et al, 1972]
Сердцевину рецептора (внутренняя колба) образуют ветвящиеся нервные терминали, окружённые пластинчатыми клетками, аналогами шванновских. Нервные терминали булавовидно расширены и содержат скопления митохондрий и везикул. Терминали не покрыты пластинчатыми клетками и отделены базальной мембраной от т.н. капсулярно-
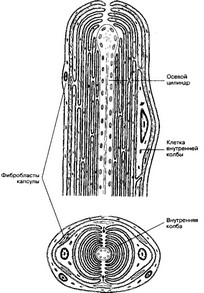
Рис. 8-15. Инкапсулированное тельце Пачини состоит из билатерально организованной внутренней колбы и наружной соединительнотканной капсулы. Клетки внутренней колбы образуют отростки, концентрическими полукольцами окружающие чувствительную нервную терминаль. Стрелками указана щель внутренней колбы, проходящая параллельно короткой оси эллипса нервной терминали [из Jun- queira LC, Carneiro J, 1991]
го пространства, расположенного между капсулой и внутренней колбой. Это достаточно обширное пространство заполнено жидкостью, содержит фибробласты, макрофаги и неориентированные коллагеновые волокна, вплетающиеся во внутреннюю колбу. Капсула тельца состоит из 4-5 слоёв уплощённых клеток.
- Тельца Пачйни (рис. 8-15) обнаружены в соединительной ткани кожи и различных органов, имеют овальную форму, размер до 0,5x1,0 мм. Внутренняя колба, наружная капсула и терминальное нервное волокно — основные компоненты тельца.
а. Внутренняя колба. Центральная часть внутренней колбы занята одиночным безми- елиновым волокном, ориентированным параллельно длинной оси тельца. Внутренняя колба состоит из нескольких десятков отростчатых клеток, образующих две разделённые щелями полуокружности. По периферии внутренней колбы расположены тела её клеток, в их цитоплазме содержатся большое количество митохондрий, хорошо развитая эндоплазматическая сеть, комплекс Гольджи, множество мелких вакуолей.
б. Наружная капсула — множество уплощённых фибробластов. Между клетками сосредоточено значительное количество коллагеновых волокон.
в. Терминаль. К тельцу Пачини подходит толстое миелинизированное нервное волокно. Внутри наружной капсулы оно образует несколько перехватов Ранвьё. Подойдя к внутренней колбе рецептора, нервное волокно теряет миелин и переходит в чувствительную нервную терминаль.
г. Дополнительное волокно (рис. 8-16). В 1896 году в Казанской гистологической лаборатории ДА Тимофеев обнаружил в тельцах Пачини дополнительное нервное волокно. Функция этого волокна неясна.
д. Функция. Адекватная механическая стимуляция телец Пачини приводит к появлению рецепторного потенциала в терминальной части нервного окончания. Достигнув критической величины, рецепторный потенциал в первом перехвате Ранвьё вызывает появление потенциалов действия. Наружная капсула рецептора — фильтр, пропускающий только динамическую составляющую механического воздействия. Рассматриваемый тип чувствительных нервных окончаний относится к фазным, или быстроадап- тирующимся рецепторам. Тельца Пачини, как и все тканевые механорецепторы, — первичночувствующие рецепторы. Это означает, что первичный трансформационный процесс происходит в нервной терминали рецептора.
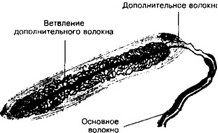
Рис. 8-16. Дополнительное волокно в инкапсулированном рецепторе. Наряду с основным волокном, в состав рецептора входит более тонкое дополнительное волокно. Терминаль основного волокна редко ветвится во внутренней колбе, тогда как дополнительное волокно образует вокруг него густую сеть [из. Тимофеев ДА, 1896]
- Тельца Майсснера (рис. 8-17) присутствуют в сосочковом слое кожи. Тельце имеет удлинённую форму, снаружи покрыто соединительнотканной капсулой. Сердцевина тельца образована пластинчатыми вспомогательными клетками, чередующимися с расширенными нервными терминалями. В формировании тельца участвует несколько миелиновых волокон.
- Другие тельца (например, колбы Краузе, генитальные тельца, тельца Голъджи-Мац- цони) встречаются реже.
- Рецепторы мышц и суставов
а. Мышечные веретёна (рис. 8-18) — чувствительные воспринимающие приборы скелетной мышцы. Их количество в различных мышцах значительно варьирует, но они присутствуют практически во всех мышцах, за исключением некоторых глазных. Основные структурные элементы мышечного веретена — интрафузальные мышечные волокна, нервные волокна и капсула.
- Мышечные волокна. Мышечное веретено содержит от I до 10 коротких интра- фузальных мышечных волокон. В средней (экваториальной) их части ядра образуют компактное скопление (волокна с ядерной сумкой) или располагаются цепочкой (волокна с ядерной цепочкой).
- Нервные волокна. Терминали Ьволокон образуют спираль в пределах экваториальной зоны обоих типов интрафузальных мышечных волокон (первичные, или
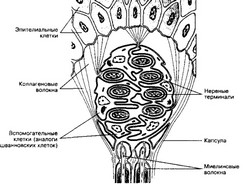
Рис. 8-17. Тельце Майсснера окружено соединительнотканной капсулой с вплетёнными в неё коллагеновыми волокнами окружающей соединительной ткани. Подходящие к тельцу нервные волокна теряют миелин и заходят внутрь тельца, где формируют многочисленные терминали, окружённые вспомогательными клетками [из Junqueira LC, CarneiroJ, 1991]
аннулоспиральные окончания). Терминали более тонких II-волокон заканчиваются на интрафузальных волокнах в области, расположенной рядом с экваториальной (вторичные окончания, чаще встречаются в волокнах с ядерной цепочкой). Эфферентные А^-волокна образуют нервно-мышечные синапсы с интрафузальны- ми волокнами в концевой их части
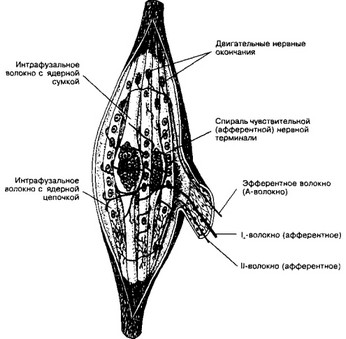
Рис. 8-18. Мышечное веретено. Интрафузальные мышечные волокна с компактным скоплением ядер — волокна с ядерной сумкой, в интрафузальных волокнах с ядерной цепочкой ядра распределены по длине волокна более равномерно. К веретену подходят афферентные и эфферентные нервные волокна. Аннулоспиральные (первичные) сенсорные окончания образованы безмиелиновыми терминалями афферентных 1о-волокон в экваториальной зоне обоих типов интрафузальных мышечных волокон Ближе к концам интрафузальных волокон (чаще волокон с ядерной цепочкой) расположены тер ми ли тонких афферентных II-волокон — вторичные окончания. Эфферентные А -волокна образуют н но-мышечные синапсы с интрафузальными волокнами в концевой их части (из Bucher О, 1980]
- Капсула. Комплекс интрафузальных волокон с нервными терминалями окружён многослойной капсулой, наружные слои которой являются производными периневрия, а внутренние рассматриваются как аналоги эндоневрия.
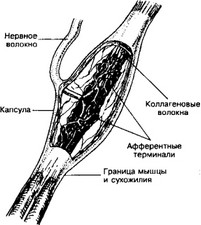
б. Сухожильные органы Гольджи (рис. 8-19) расположены в концевой части сухожилия на границе с мышцей, а также в связках капсулы суставов. Рецептор имеет веретеновидную форму и окружён капсулой, состоящей из нескольких слоёв плоских клеток. Капсула рецептора — продолжение периневрия и содержит капилляры. В образовании сухожильного органа Гольджи участвуют терминали афферентных миелиновых волокон, они ветвятся среди пучков спиралевидных коллагеновых волокон, расположенных в заполненном жидкостью пространстве.
в. Чувствительные нервные окончания в капсуле суставов — важный элемент проприоцептивной системы организма.
- Тельца Руффйни (рис. 8-14) расположены в периферических участках капсулы.
- Пластинчатые пачйниподобные тельца — сенсорные рецепторы существенно меньших размеров, чем тельца Пачйни.
- Свободные нервные окончания — терминали тонких миелиновых волокон и, наконец, терминали безмиелиновых волокон, среди которых, по-видимому, присутствуют и болевые рецепторы. Широко представлены во всех компонентах сустава, но наибольшей плотности достигают в мениске и суставном диске.
Б. Двигательные нервные окончания
-
 Нервно-мышечный синапс (рис. 8-20, см. также главу 7 I Г I). Двигательная нервная терминаль диаметром 1-1,5 мкм образует пресинаптическую область нервно-мышечного синапса, которая (в отличие от претерминальной части аксона) не содержит нейрофила- ментов и микротрубочек. В пресинаптической области в большом количестве присутствуют синаптические пузырьки и митохондрии. Для постсинаптической области характерно наличие крупных митохондрий с хорошо развитыми кристами и большого количества рибосом (существенно больше, чем в других областях саркоплазмы).
Нервно-мышечный синапс (рис. 8-20, см. также главу 7 I Г I). Двигательная нервная терминаль диаметром 1-1,5 мкм образует пресинаптическую область нервно-мышечного синапса, которая (в отличие от претерминальной части аксона) не содержит нейрофила- ментов и микротрубочек. В пресинаптической области в большом количестве присутствуют синаптические пузырьки и митохондрии. Для постсинаптической области характерно наличие крупных митохондрий с хорошо развитыми кристами и большого количества рибосом (существенно больше, чем в других областях саркоплазмы).
а. Пресинаптическая мембрана — специализированная часть аксолеммы нервной терминали. В пресинаптической мембране выявлены т.н. активные зоны — участки утолщения мембраны, имеющие прямое отношение к секреции медиатора. Рядом с активными зонами в пресинаптической мембране находятся углубления, количество которых коррелирует с уровнем секреции ацетилхолина. Пресинаптическая мембрана содержит потенциалзависимые Са2+-каналы. При деполяризации мембраны каналы открываются, и ионы Ca2+ входят в терминаль; запуская секрецию квантов ацетилхолина.
б. Синаптические пузырьки присутствуют практически в любой области нервной терминали, но в непосредственной близости от пресинаптической мембраны они образуют выраженное скопление. Размеры пузырьков варьируют, их средний
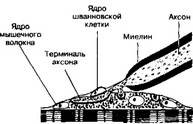
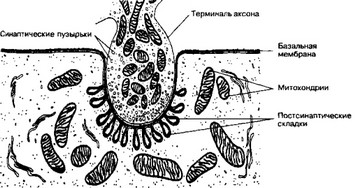
Рис. 8-20. Нервно-мышечный синапс. Пресинаптическая часть образована терминалью аксона мотонейрона и содержит скопление синаптических пузырьков вблизи пресинаптической мембраны, а также митохондрии. Постсинаптические складки увеличивают площадь поверхности постсинаптической мембраны. В синаптической щели находится синаптическая базальная мембрана (продолжение базальной мембраны мышечного волокна), она заходит в постсинаптические складки. В синаптической щели также находятся молекулы ацетилхолинэстеразы. Этот фермент расщепляет ацетилхолин и устраняет эффект деполяризующего сигнала на мышечное волокно [из Bloom W, Fawcett DW, 1968]
диаметр равен 50 нм. В холинергических синапсах светлые синаптические пузырьки содержат ацетилхолин. Часть пузырьков находится в тесном контакте с пресинаптической мембраной.
в. Постсинаптическая мембрана — специализированная часть плазмолеммы мышечного волокна — образует многочисленные инвагинации, от которых на глубину 0,5-1,0 мкм отходят постсинаптические складки, за счёт чего существенно увеличивается площадь мембраны. В постсинаптическую мембрану встроены н-холинорецепторы, их концентрация достигает 20-30 тысяч на I мкм[§§].
г. Внесинаптические холннорецепторы. Холинорецепторы присутствуют также в мембране мышечного волокна вне синапса, но здесь их концентрация на порядок меньше, чем в постсинаптической мембране.
д. Эмбриогенез и регенерация. В развивающемся мышечном волокне холинорецепторы равномерно распределены в мембране волокна. Переключение с эмбрионального на дефинитивный тип рецепторов происходит при формировании синапсов. В денерви- рованной мышце исчезают дефинитивные холинорецепторы, но чувствительность к ацетилхолину возрастает, что связано с интенсивным синтезом и включением в мембрану мышечных волокон большого количества холинорецепторов эмбрионального типа. При регенерации нервного волокна и восстановлении его контакта с мышечным волокном включается синтез холинорецепторов дефинитивного типа, встраивающихся в постсинаптическую мембрану.
е. Базальная мембрана. Через синаптическую щель проходит синаптическая базальная мембрана. Она содержитнесколько сигнальных белков (агрин, S-ламинин и др.) и играет важную роль в регуляции дифференцировки пре- и постсинаптической структуры. Синаптическая базальная мембрана удерживает в области синапса терминаль аксона, контролирует расположение холинорецепторов в виде скоплений в постсинаптической мембране. Сигнальные молекулы синаптической базальной мембраны служат метками, при помощи которых регенерирующий аксон мотонейрона находит синаптическую область на поверхности мышечного волокна.
ж. Оболочка. Снаружи нервная терминаль покрыта шванновской клеткой.
- Двигательные нервные окончания среди ГМК (см. главу 7 III).