
В пренатальном развитии человека выделяют начальный, зародышевый и плодный периоды (табл. 3-1).
Ранний эмбриогенез. Различают следующие стадии (по БВ Конюхову, 1976).
- Зигота — начало синтеза ДНК и белка.
- Дробление — начало синтеза основных типов РНК (мРНК, рРНК, тРНК).
- Морула — клетки тотипотентны.
- Бластоциста — утрата тотипотентности; клетки детерминированы к образованию зародышевых или внезародышевых структур.
Таблица 3-1. Периоды пренатального развития
|
Период |
Продолжительность периода |
Критическое событие (начало периода) |
Другие важные события периода |
|
Начальный (ранний эмбрион)* |
Первая неделя |
Оплодотворение |
|
|
Зародышевый (эмбрион) |
2-8 неделя |
Образование первичной полоски |
Органогенез Появление сердцебиений — 21-й день |
|
Плодный (плод) |
До конца беременности |
Плацентация |
|
* Считают, что эмбрион — общность клеток или существо, формирующееся на стадии первичной полоски, но не ранее. По мнению A.McLaren (1986), нельзя называть эмбрионом продукт дифференцировки зиготы до стадии первичной полоски. Автор предлагает называть этот продукт концептус, проэмбрион, предэмбрион
- Гаструла — наличие зародышевых листков и стволовых клеток.
- Органогенез — формирование зачатков органов из клеточных клонов.
- ОПЛОДОТВОРЕНИЕ
Оплодотворение — слияние мужской и женской гамет, приводящее к образованию зиготы. При оплодотворении взаимодействуют мужская и женская гаплоидные гаметы; при этом сливаются их ядра (пронуклеусы), объединяются хромосомы, и возникает первая диплоидная клетка нового организма — зигота. Начало оплодотворения — момент слияния мембран сперматозоида и яйцевой клетки, окончание оплодотворения — момент объединения материала мужского и женского пронуклеусов. Всё, что происходит до слияния мембран сперматозоида и яйцевой клетки, именуют событиями, предшествующими оплодотворению (см. III В).
А. Сперматозоид (рис. 3-24). В эякуляте человека содержится 3x10е сперматозоидов. В женских половых путях они сохраняют способность к оплодотворению максимально до 2 суток. Примерно 200 из них достигают воронки маточной трубы, где происходит встреча сперматозоида с яйцеклеткой. Сперматозоиды, не участвующие в оплодотворении, удаляются из женских половых путей или перевариваются фагоцитами.
- Направленная миграция. В феномене направленной миграции сперматозоидов известную роль играет хемотаксис. В клеточной мембране сперматозоидов существуют рецепторы к агентам хемотаксиса — N-формилпептидам.
- Капацитация. До того, как сперматозоид встретится с яйцеклеткой, он в течение нескольких часов продвигается по женским половым путям. При этом на сперматозоид воздействуют факторы женского организма (pH, слизь и др.), поддерживающие способность к миграции и оплодотворению. Этот процесс называют капацитацией.
- Простагландины, содержащиеся в эякуляте, вызывают сокращения JMK миометрия и фаллопиевых труб, облегчая продвижение сперматозоидов.
- Неподвижные сперматозоиды. При синдромах Картагенера и неподвижных ресничек сперматозоиды не передвигаются, хотя такие мужчины потенциально фертильны. В этих случаях проводят оплодотворение in vitro с последующим введением концептуса в матку.
- Дефектная акросома. По разным оценкам, эта патология составляет до 15% мужского бесплодия. Сперматозоиды имеют головку округлой формы, они подвижны, но невозможна акросомная реакция (III В 2 г)
- Азооспермия. В эякуляте нет сперматозоидов. Причины: дефекты сперматогенеза, лучевое поражение, непроходимость выводящих путей мужской половой системы.
- Олигоспермия. Уменьшенный объём эякулята (lt;1 мл) вследствие патологии яичек и придатков, семенных пузырьков, желёз (предстательная, Купера, Литтре).
Б. Яйцевая клетка (рис. 3-25). Большинство яйцеклеток в яичниках находится на стадии поздней диплотены профазы первого деления мейоза [глава 2 IV 3 а (I) (а)]. В ходе овуляции при разрыве зрелого фолликула из яичника в брюшную полость выбрасывается ово- цит второго порядка (см. главу 15.3 В 2). Его окружают прозрачная оболочка (zona pellucida) и лучистый венец (corona radiata).
- Перемещение яйцеклетки. Перед овуляцией увеличение тонуса ГМК маточной трубы приближает воронку маточной трубы к поверхности яичника. Биение ресничек эпителиальных клеток фимбрий ампулы маточной трубы создаёт ток жидкости, способствующий перемещению яйцеклетки из брюшной полости в ампулу.
При синдроме Картагенера и синдроме неподвижных ресничек женщины фертильны.
- Способность к оплодотворению. Для успешного оплодотворения яйцеклетка должна встретиться со сперматозоидом в течение одних суток после овуляции. Из практичес
ких соображений время, в течение которого овулировавшая яйцевая клетка может быть оплодотворена, оценивают в пять суток.
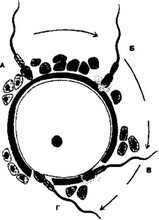
В. События, предшествующие оплодотворению. Для того, чтобы произошло оплодотворение, сперматозоид должен последовательно преодолеть три барьера (рис. 3-1): лучистый венец, состоящий из нескольких слоёв фолликулярных клеток, прозрачную оболочку и, наконец, плазматическую мембрану яйцеклетки, при слиянии которой с плазмолеммой сперматозоида и начинается собственно оплодотворение.
- Лучистый венец. Сперматозоид легко проникает через лучистый венец между рыхло расположенными фолликулярными клетками и достигает прозрачной оболочки.
- Прозрачная оболочка — существенный барьер на пути сперматозоида. При взаимодействии сперматозоида с прозрачной оболочкой последовательно происходят следующие события: связывание сперматозоида с его рецептором -gt; акросомная реакция -gt; расщепление компонентов zona pellucida ферментами акросомы -gt; проникновение сперматозоида через образовавшийся в оболочке канал к плазматической мембране яйцеклетки.
а. Строение. По мере развития яйцеклетки в ней происходят синтез и последующая секреция гликопротеинов, постепенно формирующих zona pellucida. Зрелая прозрачная оболочка содержит густую сеть тонких нитей, состоящую из гликопротеинов (главным образом, белки ZP). Zona pellucida у человека подразделяется на два слоя:
- внутренний, богатый нейтральными гликозаминогликанами;
- наружный, содержащий преимущественно кислые гликозаминогликаны.
б. Химический состав. Нейтральные и сульфатированные гликозаминогликаны, гиа- луроновая и сиаловые кислоты, гликопротеины.
-
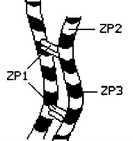
 Белки ZP. Основная масса зрелой прозрачной оболочки — гликопротеины ZP: ZPl1 ZP2 и ZP3 с M 90-110, 64-76 и 57-73 кД соответственно. ZP3 состоит из полипептида (44 кД), цепей jV-олигосахаридов и О-олигосахаридов. Синтез ZP прекращается в ходе овуляции нее или тотчас после. Прозрачная оболочка содержит примерно IO12 молекул ZP3, вместе с ZP2 формирующих нити длиной 2-3 мкм
Белки ZP. Основная масса зрелой прозрачной оболочки — гликопротеины ZP: ZPl1 ZP2 и ZP3 с M 90-110, 64-76 и 57-73 кД соответственно. ZP3 состоит из полипептида (44 кД), цепей jV-олигосахаридов и О-олигосахаридов. Синтез ZP прекращается в ходе овуляции нее или тотчас после. Прозрачная оболочка содержит примерно IO12 молекул ZP3, вместе с ZP2 формирующих нити длиной 2-3 мкм
Рис. 3-1. Преодоление сперматозоидом барьеров яйцевой клетки. А — проникновение сперматозоида через лучистый венец; Б — акросомная реакция; В — прохождение сперматозоида через прозрачную оболочку; Г — слияние клеточных мембран сперматозоида и яйцеклетки [из Alberts В et al, 1983]
и толщиной 7 нм (рис. 3-2). В составе этих нитей комплекс ZP2-ZP3 образует структурные повторы каждые 15 нм. Нерегулярным образом нити соединены при помощи ZPl1 что и приводит к образованию трёхмерной сети — каркаса прозрачной оболочки. В петлях этого каркаса расположены углеводные компоненты zona pellucida.
-

 ZP3 — рецептор сперматозоида. Инактивация или отщепление О-олигосахарида от молекулы ZP3 блокирует связывание сперматозоида с яйцеклеткой. Сперматозоид имеет рецепторы к этому олигосахариду. Главное следствие взаимодействия ZP3 прозрачной оболочки с рецептором этого гликопротеина на поверхности головки сперматозоида — начало акросомной реакции (см. Ill В 2 г).
ZP3 — рецептор сперматозоида. Инактивация или отщепление О-олигосахарида от молекулы ZP3 блокирует связывание сперматозоида с яйцеклеткой. Сперматозоид имеет рецепторы к этому олигосахариду. Главное следствие взаимодействия ZP3 прозрачной оболочки с рецептором этого гликопротеина на поверхности головки сперматозоида — начало акросомной реакции (см. Ill В 2 г). - ZP2 — вторичный рецептор сперматозоида. Как только начинается акросомная реакция, ZP2 дополнительно связывает сперматозоид.
- Блокада полиспермии. После оплодотворения ZP2 и ZP3 подвергаются химической модификации, следствием чего является блокада полиспермии (III Д 3).
в. Взаимодействие сперматозоида с прозрачной оболочкой происходит в два этапа. Сначала сперматозоид слабо прикрепляется к прозрачной оболочке. На этом этапе взаимодействие не видоспецифично, сперматозоиды легко отделяются осторожным смыванием. На втором этапе сперматозоиды прочно связываются со своими рецепторами. Это взаимодействие видоспецифично (по-видимому, видовую специфичность обеспечивает также акрозин, освобождающийся в ходе акросомной реакции).
г. Акросомная реакция — экзоцитоз содержимого акросомы для локального разрушения прозрачной оболочки и преодоления сперматозоидом этого барьера.
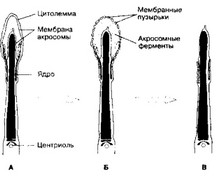
- Акросома образуется в ходе сперматогенеза как продукт комплекса Гольджи и может рассматриваться как аналог лизосом. Акросома (рис. 3-24) расположена в головке сперматозоида, кпереди от ядра и тотчас под плазматической мембраной. Спереди мембрана акросомы (наружная) соприкасается с клеточной мембраной сперматозоида, а сзади (внутренняя мембрана) — с ядерной мембраной.
- Акросомная реакция — разновидность экзоцитоза. При акросомной реакции наружная мембрана акросомы и клеточная мембрана сливаются и формируют мелкие пузырьки (рис. 3-3), отделяющиеся от головки сперматозоида. При этом из акросомы освобождаются гиалуронидаза, протеазы (в т.ч. акрозин), гли- козидазы, липазы, нейраминидаза и фосфатазы. Ферменты расщепляют молекулы прозрачной оболочки, что позволяет сперматозоиду преодолеть этот барьер.
- Механизм акросомной реакции. Взаимодействие ZP3 с цитолеммой сперматозоида приводит к массированному транспорту внутрь головки сперматозоида Ca2+ и Na* в обмен на H+. Увеличение внутриклеточной концентрации Ca2+ активирует Са2+-зависимую фосфолипазу, в результате изменяется уровень вторых посредников — циклических нуклеотидов. Вслед за этим активируется протонная
Рис. 3-3. Акросомная реакция. Последовательные этапы акросомной реакции даны на А —gt; Б —gt; В. Слияние цитолеммы сперматозоида и мембраны акросомы видно на Б [из Janagimachi R, Nada JD, 1970]
 А'ГФаза, что приводит к увеличению внутриклеточного pH. Увеличение концентрации Ca2+ в цитозоле и повышение pH в головке сперматозоида запускают акросомную реакцию.
А'ГФаза, что приводит к увеличению внутриклеточного pH. Увеличение концентрации Ca2+ в цитозоле и повышение pH в головке сперматозоида запускают акросомную реакцию.
Г. Оплодотворение. В результате акросомной реакции в прозрачной оболочке образуется узкий канал, через который проходит сперматозоид; плазматические мембраны сперматозоида и яйцеклетки вступают в соприкосновение и сливаются. Вскоре слившиеся мембраны разрушаются, головка сперматозоида оказывается погружённой в цитоплазму яйцеклетки. В течение первых 12 часов после проникновения сперматозоида в яйцеклетку происходит перестройка ядер (пронуклеусов) слившихся гамет. Ядра набухают, появляются ядрышки. Пронуклеусы мигрируют в центр яйцеклетки и сближаются. Их ядерные оболочки исчезают, а материнские и отцовские хромосомы перемешиваются — образуется синкарион (? + cf). Этот процесс (сингамия) и есть собственно оплодотворение — появляется диплоидная зигота (новый организм, пока одноклеточный). Вклад сперматозоида в оплодотворение не исчерпывается только тем, что половина хромосом диплоидной зиготы — отцовские; часть генетической информации (митохондриальный геном) новый организм получает вместе с митохондриями сперматозоида. Сперматозоид приносит также сигнальный белок дробления. Наконец, в ходе оплодотворения происходит завершение мейоза, и детерминируется генетический пол нового организма.
- Диплоидность зиготы. При сингамии (? + Cf) происходит объединение ядерного генома гаплоидных гамет: (22 аутосомы + I половая хромосома) х 2 = 44 аутосомы + 2 половые хромосомы.
- Генетический пол определяет Y-хромосома. Примерно половина сперматозоидов содержит Y-хромосому, другая половина — хромосому X. Следовательно, половина зигот имеет генетически мужской пол (XY, Cf), вторая половина — генетически женский пол (XX, ?).
а. Инактивация Х-хромосомы ($) — лайонизация (см. главу 2 II В I).
б. Детерминация пола (см. главы 15.1 Б 2 а и 18).
в. Гермафродитизм (см. главу 18).
г. Трисомия (см. главу I E 2 6).
- Завершение мейоза. Оплодотворённый овоцит второго порядка завершает деления мейоза; образуется второе полярное тельце, располагающееся рядом с первым между прозрачной оболочкой и плазмолеммой зиготы, в перивителлиновом пространстве,
формирующемся при оплодотворении в результате уменьшения объёма зиготы (см. III Д I а).
Иногда второе полярное тельце не выталкивается из зиготы (более того, возможно вхождение его хромосом в состав синкариона — самооплодотворение), в этом случае в яичниках будущего организма может развиться овариальная тератома.
- Митохондриальный геном. Самореплицирующиеся митохондрии имеют собственную ДНК (митохондриальная ДНК), кодирующую полипептиды, частично обеспечивающие функцию этого органоида. Митохондрии зиготы имеют двоякое происхождение: часть содержалась в овоците (?, материнская часть митохондриального генома), другую часть этих органоидов приносит сперматозоид (d\ отцовский митохондриальный геном). Известны наследуемые с митохондриями (отцовскими или материнскими) заболевания (например, лёберовская атрофия сетчатки, MERRF- и MELAS-синдромы, дилатационная митохондриальная кардиомиопатия, некоторые миопатии).
- Сигнальный белок дробления. В зиготе обнаружены два принесённые сперматозоидом белка с Mr 14 и 18 кД, содержащие одну и ту же Аг-детерминанту. AT к этому Ar блокируют первые дробления зиготы, не оказывая влияния на другие процессы.
Д. События после оплодотворения. В ходе собственно оплодотворения и сразу после него в зиготе и окружающих её структурах происходит ряд процессов (уменьшение объёма зиготы, формирование перивителлинового пространства, кортикальная реакция, модификация прозрачной оболочки). Назначение этих процессов — предупреждение полиспермии и создание гомеостатической среды для концептуса.
- Зигота. При слиянии плазматических мембран гамет и объединении ядерных геномов происходят существенные изменения внутриклеточного ионного состава, приводящие к уменьшению объёма зиготы, деполяризации её плазмолеммы, развитию кортикальной реакции.
а. Уменьшение объёма. Прямое следствие этого процесса — образование перивителлинового пространства. Перивителлиновое пространство содержит оба полярных тельца и имеет характерный ионный состав (гомеостатическая среда для концептуса), создающий дополнительное препятствие для проникновения к зиготе других сперматозоидов.
б. Деполяризация плазматической мембраны предупреждает проникновение сперматозоидов в зиготу.
в. Кортикальная реакция. По периферии яйцеклетки расположены кортикальные гранулы. Их диаметр 200-600 нм; они содержат ферменты, в т.ч. различные гидро- лазы. Сразу после проникновения сперматозоида в яйцеклетку начинается кортикальная реакция — выделение содержимого кортикальных гранул в перивителлиновое пространство. Сигнал для кортикальной реакции — повышение концентрации Ca2+ в цитозоле. Под действием ферментов кортикальных гранул происходят протеолиз ZP2 (ZP2 —gt; ZP2f) и модификация рецептора сперматозоида ZP3 (ZP3 —gt; ZP3f). В модифицированном состоянии ZP3f теряет способность к запуску акросом- ной реакции. Тем самым блокируется полиспермия.
- Прозрачная оболочка. В результате кортикальной реакции прозрачная оболочка подвергается существенным изменениям — происходит её стабилизация (оболочка оплодотворения). В стабилизированном состоянии прозрачная оболочка защищает проходящий по маточной трубе концептус. Без прозрачной оболочки дробление зиготы невозможно.
- Блокирование полиспермии
а. Деполяризация мембраны зиготы (III Д I б).
б. Перивителлиновое пространство (III Д I а).
Внутриутробное развитие 79
в. Стабилизация прозрачной оболочки (III Д 2).
г. Блокада акросомной реакции (III Д I в).
Е. Экстракорпоральное оплодотворение. В акушерско-гинекологической практике часто сталкиваются с невозможностью иметь ребёнка вследствие патологии маточных труб, оли- госпермии и т.д. Проблему решают применением экстракорпорального оплодотворения. Сущность метода состоит в получении жизнеспособных яйцеклеток с последующим их оплодотворением in vitro, введением в матку концептуса и возникновением беременности. Если в естественных условиях при овуляции из яичника выходит одна яйцеклетка, то для успешного экстракорпорального оплодотворения необходимо одновременное получение большого их количества. Для этого женщине проводят медикаментозную стимуляцию овуляции гормональными препаратами. Яйцеклетки получают при хирургической операции с применением эндоскопического оборудования или при помощи трансвагинальной техники. Полученные яйцеклетки культивируют in vitro в питательной среде специального состава, где и производят их оплодотворение путём введения в среду сперматозоидов. Дробление контролируют микроскопически, и 4 нормально развивающихся концептуса одновременно вводят через шейку в полость матки. Операция успешна в 20-25% случаев.