
Воспроизведение возможных филогенетических событий, обеспечивших формирование суперсемейства иммуноглобулинов и в первую очередь антигенраспознающих структур, должно включать три определяющих положения:
- в основе происхождения всех членов суперсемейства лежал предковый однодоменный пептид;
- локусы, которые контролируют иммуноглобулинподоб- ные молекулы, включающие несколько (от двух до семи) доменов, возникли в результате тандемных дубликаций исходного гена для однодоменного пептида;
- разные домены суперсемейства обладают одним из двух способов взаимодействия между собой — гомофильной и гетерофильной формой.
Устоялось мнение, что гетерофильной форме взаимодействия предшествовали гомофильные молекулярные отношения. Цепь событий, приведших к формированию антигенраспознающих рецепторов как завершающих молекулярных структур в эволюции суперсемейства, представляется следующим образом (рис. 5.2). Пред- ковые однодоменные белки, связанные с поверхностью клетки, в процессе межклеточного взаимодействия формировали гомодимер (A-А). Подобная форма отношений, очевидно, имела место при возникновении многоклеточных организмов. Гомофильная форма взаимодействия сохранилась до настоящего времени у однодоменного белка Р0 и, возможно, Thy-1. В результате тандемной дупликации гена для однодоменного белка и последующих мутационных изменений, затрагивающих контактирующий участок молекулы, стали проявляться гетерофильные клеточные отношения (А- В). Дуплицированные гены экспрессировались в клетках одного и того же или разных типов. Мутационные изменения могли снижать аффинность А-В-взаимодействий, но не отменять их полностью. Возникшие гетерофильные отношения явились источником формирования V-доменов Т-клеточного рецептора (от домена В) и антигенпрезентирующей способности молекул МНС (от домена А).
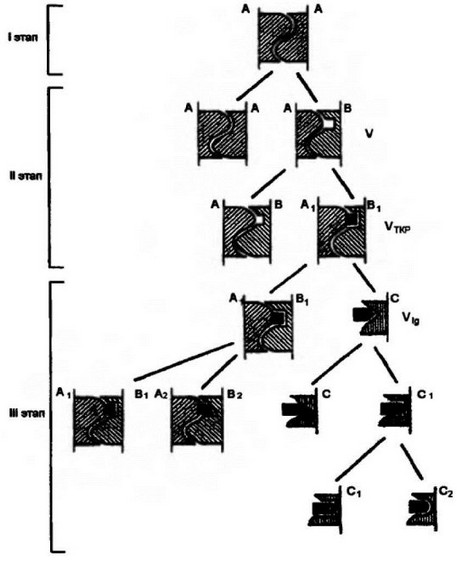
Рис. 5.2. Схема возможных путей вознижвовеянж специфических Т-клеточных, шгтигенрасооттющнх рецепторов (ТКР), поверхностных иммуноглобулиновых рецепторов (slg) и антител.
А-Ад — молекулы МНС и их комплекс с антигенными пептидами, В-Вг — вариабельные домены ТКР (Vtkp)* С-С2 — вариабельные домены slg и антител
(V|g); А-А — гомофильное взаимодействие, А-В А2-В2 — гетерофильные
взаимодействия
Мутационные изменения в домене В носили множественный характер, но среди этих изменений были и такие, которые создавали условия для взаимодействия с комплексом А+чужеродный антигенный пептид. В качестве чужеродных пептидов могли выступать вирусы или измененные собственные антигены. В такой ситуации V-ген Т-клеточного рецептора (Vncp-ген) попадал под положительное давление отбора. Последующие множественные дупликации Уткр-гена и параллельные и независимые мутационные изме-
нения этих вновь возникших генов поставляли материал для дальнейшего действия естественного отбора, что и создавало набор генов для V-доменов Т-клеточных рецепторов, способных взаимодействовать с множеством различных чужеродных пептидов, комплексированных с антигенами МНС. В свою очередь гены для доменов МНС также эволюционировали, создавая полиморфизм и полигению молекул этого комплекса. Однако эволюция как молекул МНС, так и Т-клеточных рецепторов имела свои ограничения. Широкая изменчивость касалась только участков доменов, непосредственно взаимодействующих с антигенным пептидом или распознающих этот пептид. В значительно меньшей степени изменения затрагивали те участки, которые вступают в контактные отношения независимо от антигена (подробнее см. главу 3).
Т-клеточные рецепторы эволюционно предшествовали иммуноглобулинам, и V-гкр-гены были исходными для Уц-генов. Об этом, в частности, говорят факты эволюционно более раннего проявления Т-клеточного иммунитета по сравнению с гуморальным иммунитетом (см. главу 20).
Качественное отличие поверхностного иммуноглобулина В- клеток от Т-клеточного рецептора состоит в том, что антигенрас- познающий рецептор В-клеток взаимодействует собственно с экзогенным пептидом без включения продуктов МНС в качестве распознаваемых структур. Кроме того, аффинность В-клеточного рецептора к антигенам значительно выше таковой Т-клеточного рецептора. Константа диссоциации (KD) для Т-клеточного рецептора составляет 10"М05, в то время как этот показатель для поверхностного иммуноглобулина и антител находится в пределах 106- 10,0. Вероятно, эволюция иммуноглобулинов пошла по линии освобождения зависимости от дополнительных клеточных образований (молекул МНС, корецепторов) и компенсаторного, через усиление аффинности, взаимодействия с экзогенным свободным антигеном.
Эволюция Cl-доменов на самых начальных этапах проходила, очевидно, самостоятельно, с умеренными дупликациями и мутационными изменениями. Факторами отбора в данном случае были не антигены, а иные, физиологические механизмы. У позвоночных животных в результате транслокационных перестроек генов для V- и С-доменов возник единый информационный участок, контролирующий синтез различных изотипов иммуноглобулинов.
Эволюционные изменения С2-доменов, входящих в структуру адгезивных и рецепторных молекул, остаются непонятыми, даже в гипотетическом плане.
Одно из существенных свойств эволюции суперсемейства иммуноглобулинов, как, впрочем, и дрgt;гих систем иммунитета,
состоит в том, что в процессе филогенетического развития этого суперсемейства появляющиеся вновь функционально более совершенные молекулы не исключали предыдущих, от которых они возникли. Так, например, предковые домены Thy-1, р2-м, Ро, стоявшие у истоков формирования суперсемейства, широко представлены у современных видов. Возникновение доменов иммуноглобулиновых рецепторов и антител от доменов Т-клеточных рецепторов не отменило значимую, развившуюся функцию этих последних структур.
Заключение
Функциональное предназначение суперсемейства иммуноглобулинов разнообразно. Однако основное определяющее содержание системы — это обеспечение эффективного распознавания антигена с помощью двух специализированных классов молекул — иммуноглобулинов и Т-клеточных рецепторов.
Эталонной молекулой суперсемейства является, конечно, иммуноглобулин. Собственно именно эта молекула дала название всему суперсемейству. Тот или иной белок может быть включен в состав данного молекулярного объединения при соблюдении двух условий — соответствия пространственной организации своих доменов особенностям конформации доменов иммуноглобулинов и достаточной степени гомологии с последовательностью аминокислот в доменах иммуноглобулинов.
При распознавании ассоциированного с клеткой антигена Т-клеточный рецептор действует не изолированно, а с помощью дополнительных молекул — CD3 y,S,e, CD4 и CD8. С процессом антигенного распознавания связаны также молекулы I и II классов МНС, выполняющие функцию представления антигена в иммуногенной форме для его распознавания Т-клеточными рецепторами.
Адгезивные и рецепторные молекулы суперсемейства: LFA3, CD2, Poly-IgR, Fcy2b/ylR, CSF 1R и др., — не принимают участия в процессе антигенного распознавания, но являются активными участниками регуляции иммунного ответа. Так, CD2 и LFA3 способствуют установлению контакта между Т-клетками и анти- генпрезентирующими клетками, Poly-IgR обеспечивает проникновение IgM и IgA через эпителиальные барьеры, CSF 1R способствует дифференцировке клеток моноцитарного ряда.
Функция других белков суперсемейства (Р0, Thy-1, LINK, а 1В), в иммунном реагировании неизвестна. Однако в отношении однодоменных белков ясно другое: именно они явились предко- выми формами для эволюционного формирования всего суперсемейства.
Эволюция суперсемейства иммуноглобулинов была сопряжена с эволюцией иммунокомпетентных клеток, что приводило ко все большему филогенетическому совершенствованию системы специфической защиты в целом.