Рецепторы для цитокинов
Действие цитокинов осуществляется через рецепторы. По особенностям структуры полипептидных цепей выделяют несколько групп цитокиновых рецепторов. Приводимую классификацию применяют именно к полипептидным цепям. В состав одного рецептора могут входить цепи, относящиеся к разным семействам. Важность этой классификации обусловлена тем, что для разных типов полипептидных цепей рецепторов характерен определенный сигнальный аппарат, состоящий из тирозинкиназ, адапторных белков и транскрипционных факторов.
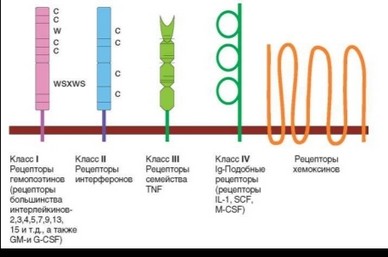
В табл. 2.29 представлено 6 структурных вариантов цитокиновых рецепторов, из которых 5 отображено на рис. 2.50. Наиболее многочисленный тип — цитокиновые гемопоэтиновые рецепторы. Для их внеклеточных доменов характерно наличие 4 остатков цистеина и присутствие последовательности, содержащей остатки триптофана и серина — WSXWS. Домены семейства фибронектина, содержащие 4 остатка цистеина, составляют основу рецепторов интерферонов. Характерная черта доменов, образующих внеклеточную часть рецепторов семейства TNFR, — высокое содержание остатков цистеина («богатые цистеином домены»). Эти домены содержат 6 остатков цистеина. Группа рецепторов, внеклеточные домены которых относят к суперсемейству иммуноглобулинов, включает две группы — рецепторы для IL-1 и несколько рецепторов, цитоплазматическая часть которых обладает тирозинкиназной активностью. Тирозинкиназная активность свойственна цитоплазматической части практически всех ростовых факторов (EGF, PDGF, FGF и т.д.). Наконец, особую группу образуют родопсиноподобные рецепторы хемокинов, 7-кратно пронизывающие мембрану, которые были описаны выше (см. раздел 2.3.2.2). Однако не все полипептидные цепи рецепторов соответствуют этой классификации. Так, ни a-, ни р-цепи рецептора IL-2 не относят к семействам, представленным в табл. 2.29 (a-цепь содержит домены контроля комплемента). В основные группы также не входят рецепторы IL-12, общая p-цепь рецепторов IL-3, IL-5, GM- CSF и некоторые другие полипептидные цепи рецепторов.
2.5. Гуморальные факторы врожденного иммунитета
Таблица 2.29. Классификация цитокиновых рецепторов
|
Название групп рецепторов |
Число внеклеточных доменов |
Характеристика |
Цитокины- лиганды |
|
I. Гемопоэтиновые рецепторы |
2-7 |
4 остатка Cys и наличие последовательности WSXWS |
IL-2, IL-3, IL-4, IL-5, IL-6, IL-7, IL-8, G-CSF и GM-CSF, EPO |
|
II. Рецепторы интер- феронов |
1-2 |
4 остатка Cys |
IFNa, IFNp, IFNy, IL-10 |
|
III. Рецепторы фактора некроза опухоли |
1 |
При взаимодействии с лигандами образуют тример |
TNFa , лимфотоксин a |
|
IV. Рецепторы интерлейкина-1 |
3 |
Внеклеточные домены суперсемейства иммуноглобулинов, внутриклеточный (TIR) сходен с TIR доменом TLR |
IL-1, IL-18 |
|
V. Иммуноглобулиноподобные рецепторы |
5 |
Внеклеточные домены — суперсемейства иммуноглобулинов, внутриклеточные — тирозинкиназы |
M-CSF, SCF (c-kitL), Flt3L |
|
VI. Рецепторы хемо- кинов |
Внеклеточные домены отсутствуют |
Родопсиноподобные рецепторы, 7-кратно пронизывающие мембрану. Связаны с G-белком |
Хемокины |
|
VII. Другие |
Обычно 1 |
Различные типы структуры |
IL-2, IL-15, TGFp |

Рис. 2.51. Разновидности рецепторов для IL-2. Связь аффинности с субъединичным составом. Указаны цепи (а, р, у), лиганд (IL-2) и Jak-киназы, связанные с полипептидными цепями
Практически все цитокиновые рецепторы (кроме иммуноглобулиноподобных, обладающих киназной активностью) состоят из нескольких полипептидных цепей. Нередко разные рецепторы содержат общие цепи. Наиболее яркий пример — у-цепь, общая для рецепторов IL-2, IL-4, IL-7, IL-9, IL-15, IL-21, обозначаемая как у(с). Дефекты этой цепи играют важную роль в развитии иммунодефицитной патологии (см. раздел 4.7.1.5). Общая p-цепь входит в состав рецепторов GM-CSF, IL-3 и IL-5. Общие цепи имеют IL-7 и TSLP (а-цепь), а также IL-2 и IL-15, IL-4 и IL-13 (в обоих случаях — р-цепь).
Роль субъединичного состава в формировании аффинности рецептора удобно иллюстрировать на примере рецептора для IL-2. Все полипептидные цепи, входящие в его состав, участвуют в формировании связывающего участка для IL-2 — 3 варианта этого рецептора с разным сродством к IL-2 (низким, промежуточным и высоким) различаются по «степени укомплектованности» полипептидными цепями (рис. 2.51). Так, отдельно экспрессируемая а-цепь способна связывать IL-2 с низким сродством (И=10-8 М), но не способна передавать сигнал в клетку. Димер Ру обладает промежуточным сродством к IL-2 (Kd=10-9 M); этот вариант рецептора присутствует на покоящихся клетках и естественных киллерах, причем на последних он передает сигнал. Активированные Т-клетки, а также регуляторные Т-клетки и некоторые естественные киллеры экспрессируют высокоаффинный тримерный рецептор состава аРу с Kd=10-10 М.
Как правило, рецепторы представлены на поверхности покоящихся клеток в небольшом количестве и нередко в неполном субъединичном составе. Обычно в таком состоянии рецепторы обеспечивают адекватный ответ только при действии очень высоких доз цитокинов. При активации клеток число мембранных рецепторов цитокинов увеличивается на порядки, более того, эти рецепторы «доукомплектовываются» полипептидными цепями, как это было показано выше на примере рецептора для IL-2. Под влиянием активации число молекул этого рецептора значительно возрастает и в их составе появляется а-цепь, ген которой экспрессируется в процессе акти
вации. Благодаря таким изменениям лимфоцит приобретает способность пролиферировать в ответ на действие IL-2.
Источник: Ярилин.А.А , «Иммунология » 2010