
Гемолиз (от греческого слова Haima - кровь, Lysis - разрушение) - разрушение клеток гемопоэза. Продолжительность жизни эритроцитов составляет от ПО до 130 дней, в среднем 120 дней. В течение одной минуты эритроцит дважды проходит через капилляры меньшего диаметра (2-4 мкм), чем диаметр эритроцита (большая часть которых имеет d 7,2-7,5 мкм). За период жизни эритроцит покрывает расстояние в 150 200 км, из которых около половины составляют узкие территории. На некоторое время эритроциты застаиваются в синусах селезенки, где сосредоточена специализированная система фильтра и удаления состарившихся эритроцитов.
Внутриклеточный гемолиз
В нормальном организме существует постоянное равновесие между продукцией и деструкцией кроветворных клеток. Основная масса эритроцитов разрушается путем фрагментации (эритрорексиса) с последующим лизисом и эритрофагоцитозом в органах ретикулоэндотелиальной системы (РЭС), преимущественно в селезенке, частично в печени. Нормальный эритроцит проходит синусы селезенки благодаря своему свойству изменять форму. По мере старения эритроциты теряют способность деформироваться, задерживаются в синусах селезенки и секвестрируются.
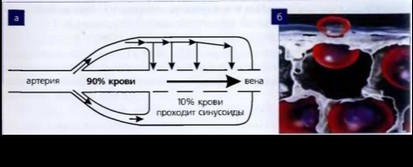
Из поступившей в селезенку крови 90% эритроцитов проходит, не задерживаясь и не подвергаясь фильтрационному отбору (рис. 68).
10% эритроцитов попадает в систему сосудистых синусов и вынуждены выбираться из них, профильтровываясь через поры (фенестры), размер которых на порядок меньше (0,5-0,7 мкм), чем диаметр эритроцита. У старых эритроцитов изменена ригидность мембраны, они застаиваются в синусоидах. В синусах селезенки снижен pH и концентрация глюкозы, поэтому
при задержке в них эритроцитов последние подвергаются метаболическому истощению. Макрофаги расположены по обеим сторонам синусов, их основная функция - элиминировать старые эритроциты. В макрофагах РЭС заканчивается разрушение эритроцита (внутриклеточный гемолиз). В нормальном организме с помощью внутриклеточного гемолиза разрушается почти 90% эритроцитов.
Механизм распада гемоглобина в клетках РЭС начинается с одновременного отщепления от него молекулы глобина и железа. В оставшемся тетрапиррольном кольце под действием фермента гемоксигеназы происходит окисление а-метиновой связи, при этом гем теряет свою цикличность, образуя линейную структуру - биливердин. На следующем этапе путем ферментативного восстановления у-метиновой связи биливердин-редуктазой биливердин превращается в билирубин. Билирубин, образованный в РЭС, поступает в кровь, связывается с альбумином плазмы и в таком комплексе поглощается гепатоцитами, которые обладают селективной способностью захватывать билирубин из плазмы. До поступления в гепатоцит билирубин носит название неконъюгированный, или непрямой. При высокой гиперби- лирубинемии небольшая часть может оставаться несвязанной с альбумином и фильтроваться в почках.
Паренхиматозные клетки печени адсорбируют билирубин из плазмы с помощью транспортных систем белков мембраны гепатоцита - протеина Y (лигандин) и протеина Z. Место пребывания протеина Z в основном в слизистой кишечника, однако при высокой билирубинемии после насыщения протеина Y он включается в транспорт пигмента и в печени. В гепатоците неконъюгированный билирубин подвергается конъюгации главным образом с глюкуроновой кислотой. Этот процесс катализируется ферментом уридилдифосфат(УДФ)-глюкуронилтрансферазой с образованием конъюгированного билирубина в виде моно- и диглюкуронидов. Активность фермента снижается при поражении гепатоцита. Активность этого фермента, так же как и лигандина, низкая у новорожденных. Поэтому печень новорожденного не в состоянии переработать больших количеств билирубина распадающихся избыточных эритроцитов, и у новорожденных развивается физиологическая желтуха.
Конъюгированный билирубин выделяется из гепатоцита с желчью в виде комплексов с фосфолипидами, холестерином и солями желчных кислот. Дальнейшее преобразование билирубина происходит в желчных путях под влиянием дегидрогеназ с образованием уробилиногенов, мезобилирубина и Других производных билирубина. Уробилиноген в двенадцатиперстной кишке всасывается энтероцитом и с током крови воротной вены возврашается в печень, где окисляется до конечных продуктов - дипирролов.
Остальной билирубин и его производные поступают в кишечник, в котором последовательно восстанавливаются (3- и а-метиновые связи, и превращаются в стеркобилиноген. Основная масса стеркобилиногена в толстой кишке подвергается окислению в стеркобилин и выделяется с калом. Небольшая часть всасывается в кровь и через геморроидальные вены и нижнюю полую вену поступает в почки и выводится с мочой.
Следовательно, билирубин экскретируется из организма в виде стерко- билина кала и уробилина мочи. По концентрации стеркобилина в кале можно судить об интенсивности гемолиза. От концентрации стеркобилина в кишечнике зависит и степень уробилинурии. Однако генез уро- билинурии определяется также функциональной способностью печени к окислению уробилиногена. Поэтому увеличение уробилина в моче может свидетельствовать не только о повышенном распаде эритроцитов, но и о поражении гепатоцитов.
Лабораторными признаками повышенного внутриклеточного гемолиза являются: увеличение содержания в крови неконъюгированного билирубина, стеркобилина кала и уробилина мочи.
Патологический внутриклеточный гемолиз может возникнуть вследствие:
- наследственной неполноценности мембраны эритроцита (эритроци- топатии);
- нарушения синтеза гемоглобина и ферментов (гемоглобинопатии, энзимопатии);
- изоиммунологического конфликта по групповой и Rh-принадлежности крови матери и плода;
- избыточного количества эритроцитов (физиологическая желтуха, эритробластоз новорожденного, эритремия - при количестве эритроцитов более 6-7 х 1012/л).