
К скотобактериям (Scotobacteria) относятся индифферентные к свету грамотрицательные, не образующие спор бактерии. По морфологии организмов и разнообразию физиологических групп это самый обширный класс в царстве Procaryotae. Он включает все виды хемолитотрофных бактерий, близких к ним по физиологии - метилотрофных и значительную часть хемогетеротрофных видов.
Хемодитотрофные бактерии получают необходимую для жизнедеятельности энергии за счет окисления неорганических субстратов. В зависисмости от окисляемого субстрата они делятся на несколько физиологических групп (табл.9).
Таблица 9
Физиологические группы хемолитогрофных бактерий
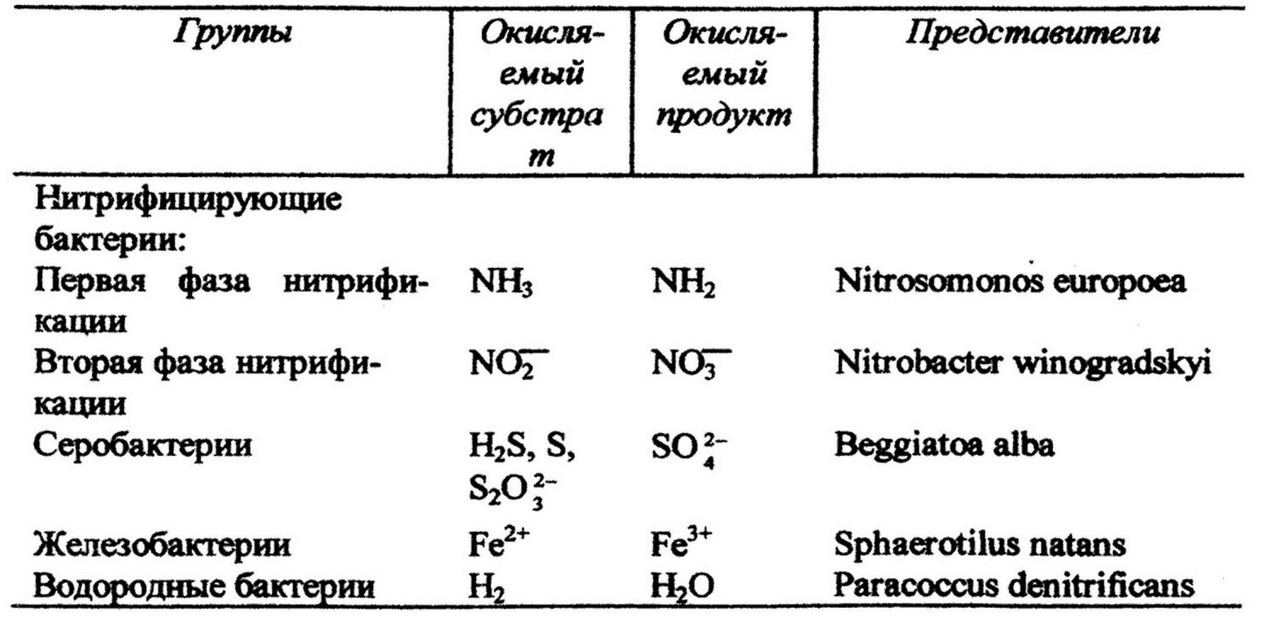
Хемолитотрофные бактерии разнообразны по морфологии и физиологии, широко распространены в природе и играют ведущую роль в круговороте биогенных элементов.
Метилотрофные бактерии получают энергию и углерод в ходе метаболизма одноуглеродных соединений, содержащих метильную группу. Это типичные облигатные метилотрофы. Наряду с ними существуют и факультативные метилотрофы, которые кроме С 1-соединений используют также С2-С6-соединения (табл. 10).
Таблица 10
Метилотрофные бактерии
|
Группа |
Род |
Форма клеток |
Метабол из ируе- мые соединения углерода |
|
Облигатные |
Methylosinus |
Палочки с полярно |
Метан, |
|
метилотрофы |
|
расположенными |
диметиловый |
|
|
|
жгутиками; |
эфир, метиловый |
|
|
|
образуют экзоспоры |
спирт |
|
|
Methylocystis |
Неподвижные |
|
|
|
|
палочки, экзоспор |
|
|
|
|
не образуют |
|
|
|
Methylomonas |
Палочки с полярно |
|
|
|
|
расположенными |
|
|
|
|
жгутиками |
|
|
|
Methyl obacter |
Палочки и кокки, |
|
|
|
|
образуют цисты |
|
|
|
Methylococcus |
Неподвижные кокки |
|
|
Факультатив |
Hyphomicro- |
Палочки с заострен |
Метиловый, эти |
|
ные метило |
bium |
ными концами, об |
ловый спирты; |
|
трофы |
|
разуют гифы и |
муравьиная кис |
|
|
|
почки |
лота, уксусная и |
|
|
Pseudomonas |
Палочки с полярно |
Р-оксим асляная |
|
|
(не все виды) |
расположенными |
кислоты, метил |
|
|
|
жгутиками |
амины |
|
|
Mycobacterium |
Палочки, иногда |
|
Как видно из таблицы 10, круг соединений,
метаболизируемых метилотрофами, невелик.
Род Pseudomonas объединяет палочковидные, неспорообразующие подвижные бактерии с полярно расположенными жгутиками. Клетки имеют вид прямых или слегка изогнутых палочек размером 1,0-0,5 х 1-4 мкм. В большинстве своем - строгие аэробы, развиваются в широком диапазоне температур - от 4 до 43° С. Многие псевдомонады не нуждаются в факторах роста. Они обладают редкой способностью использовать широкий круг источников питания - до 150 наименований природных и синтетических соединений. Уникальной особенностью псевдомонад является способность потреблять ароматические соединения, такие, как бензоат, фенол, нафталин, камфору, салицилат, не утилизируемые другими бактериями. Они могут также разлагать некоторые токсические соединения, в том числе пестициды, содержащие в своем составе хлор, фтор, ртуть. Некоторые псевдомонады, как Ps. saccharophila, Ps. facilis, являются факультативными хемолитотрофами, в качестве источника энергии используют водород или углекислый газ, другие - способны к денитрификации, т. е. добывают энергию путем восстановления нитратов, например, Ps. denitrificans, Ps. stutzeri.
Характерным свойством псевдомонад является утилизация сахаров по пути Энтнера-Дудорова, т. е. через образование 2-кето-З- дезокси-6-фосфоглюконовой кислоты (КДФГ-путь). Среди прокариот псевдомонады являются уникальными гетеротрофами, так как неизвестно ни одного природного или полученного путем химического синтеза соединения, которое не потреблялось бы этими бактериями. Наряду с потреблением псевдомонады способны и к биосинтезу различных соединений. Высокой биосинтетической способностью отличаются флуоресцирующие псевдомонады: Ps. fluorescens, Ps. aeruginosa, Ps. putida. Многие виды синтезируют ряд (более 30) антибиотиков активных в отношении грибов, грамположительных и грамотрицательных бактерий. По способности продуцировать эти биологически активные вещества они уступают только актиномицетам. Практическое применение уже получил антибиотик пирролнитрин, продуцируемый разными
видами бактерий рода Pseudomonas (Ps. purrocinia, Ps. aureofaciens. Ps. acidula и др.).
Пирролнитрин обладает высокой антифунгальной активностью, а также оказывает бактерицидное действие на сальмонеллы, кишечную палочку, золотистого стафилококка и др. Применяется для лечения дерматомикозов. В 1976 г. из Ps. sorbistini nov. sp. выделен комплекс антибиотиков аминогликозидов - сорбистины. Они характеризуются широким спектром действия: подавляют рост грамположительных и грамотрицательных бактерий, в том числе бактерий, резистентных к другим аминогликозидам.
Почти все представители рода Pseudomonas обладают способностью к синтезу бактериоцинов. Большой интерес представляют псевдомонады для технической микробиологии и биотехнологии как продуценты органических кислот, аминокислот, ферментов. Ряд штаммов уже используется для получения пировиноградной, глюконовой, а-кетоглутаровой кислот, глутаминовой и аспарагиновой аминокислот, гидролитических ферментов - липазы, эстеразы и др. Псевдомонады заслуживают внимания и как продуценты белка на одноуглеродных соединениях - метане, метаноле, формальдегиде, которые они могут использовать как единственные источники углерода и энергии.
Бактерии рода Pseudomonas - обширная группа микроорганизмов, которые повсеместно распространены в природе: почве, морских и пресных водах, илах, сточных водах. Среди них много сапрофитов, но есть также виды, патогенные для человека и животных (Ps. aeruginosa, Ps. mallei) и для растений (Ps. syringae, Ps. solanacearum).
Фенотипическое сходство с бактериями рода Pseudomonas имеют бактерии рода Xanthomonas. Характерным признаком бактерий рода Xanthomonas является образование желтых внутриклеточных пигментов - бромированных производных арилоктанов, не обнаруженных у других видов бактерий. Входящие в состав рода Xanthomonas бактерии являются патогенными для растений.
Род Azotobacter составляют крупные подвижные клетки, склонные к изменению морфологии от палочковидной до кокковидной в зависимости от возраста культуры. Для азотобактера характерно образование покоящихся форм цист, которые устойчивы к высушиванию и обеспечивают выживание азотобактера в почве в периоды ее пересыхания. Азотобактер - свободноживущий азотфиксатор, который в отличие от всех других азотфиксирующих бактерий производит азотфиксацию в аэробных условиях. Азотфиксирующий ферментный комплекс нитрогенеза защищается от токсического действия молекулярного кислорода благодаря высокой интенсивности дыхания.
С азотобактером сходны бактерии рода Azomonas, но они не образуют цисты и основным местообитанием их является вода.
Бактерии рода Beijerinckia распространены в тропических кислых почвах. Они также не способны к образованию цист.
Род Rhizobium (клубеньковые бактерии) - подвижные, не образующие спор палочковидные бактерии. Обитают в почве или корневых клубеньках бобовых растений, с которыми у них сложились симбиотические взаимоотношения. Находясь в клубеньках растений, они активно фиксируют молекулярный азот. В свободном состоянии (в почве или питательной среде)
азотфиксирующая способность клубеньковых бактерий выражена слабо. Для проявления ее необходимо обеспечить эти бактерии подходящими источниками углерода, преимущественно пентозами, промежуточными соединениями цикла Кребса и минимальными количествами связанного азота. До 1975 г. вообще отрицалась способность клубеньковых бактерий в чистой культуре связывать молекулярный азот. В качестве источника углерода и энергии клубеньковые бактерии потребляют широкий круг органических соединений: углеводы, спирты, органические кислоты и их соли. Источником азота, кроме молекулярного, служат нитраты, соли аммония, аминокислоты, азотистые основания. В зависимости от условий аэрации нитраты могут служить не только источником азота, но и акцептором электронов. В анаэробных условиях большинство видов клубеньковых бактерий осуществляют редукцию нитратов, только R. lupini не обладают нитратредуктазной активностью. Классификацию клубеньковых бактерий производят по растению-хозяину, на котором бактерии образуют клубеньки.
Клубеньковые бактерии - аэробы, но способны расти при пониженном парциальном давлении кислорода (менее 0,01 атм) Оптимальная температура роста 25-30° С, пределы pH 5,0-8,5.
Отношения между клубеньковыми бактериями и бобовыми растениями определяются как мутуализм, когда оба симбионта извлекают выгоду из сожительства: растение получает азот, клубеньковые бактерии - углеродсодержащие вещества и минеральные соли.
Бактерии рода Agrobacterium по морфологическим н культуральным свойствам сходны с клубеньковыми бактериями. Но s отличие от последних агробактерии не способны фиксировать молекулярный азот, вызывают образование галлов, или опухолей на стеблях и корнях растений разных семейств. Типичный представитель рода A. tumefaciens индуцирует образование корончатых галлов более чем у 40 семейств. Бактерии внедряются в ткань растений через повреждения и вызывают разрастание стебля.
Род Caulobaster включает бактерии оригинальной формы с уникальным жизненным циклом. Эти бактерии образуют нитевидные клеточные выросты - простеки, которые представляют собой выпячивание клеточного содержимого, соединенного с цитоплазмой клетки. На конце выроста содержится клейкое вещество, с помощью которого каулобактер прикрепляется к твердому субстрату или клеткам других микроорганизмов Клетки каулобактер а имеют палочковидную форму с одним полярно расположенным жгутиком. Размножаются бинарным делением. При этом образуются две неодинаковые дочерние клетки: одна из клеток несет простеку (стебелек), другая - жгутик. Клетка, несущая простеку, неподвижна; сразу после деления на противоположном простеке полюсе формируется жгутик, и клетка снова может делиться таким же способом. Клетка, снабженная жгутиком, претерпевает морфологические преобразования: теряет жгутик и на его месте образует стебелек. Затем прикрепляется к субстрату и переходит в вегетативную фазу.
Бактерии рода Asticcacauiis отличаются от бактерий рода Caulobacter субполярным расположением жгутика и простеки Стебельковые бактерии группы каулобактера встречаются как в пресной, так и в морской воде. Развиваются в основном за счет
потребления веществ, секретируемых другими микроорганизмами, к которым они прикреплены. В торфяниках и илах обнаружены бактерии звездообразной формы, получившие название Stella gumosa.
Спирохеты. Это сравнительно небольшая группа одноклеточных микроорганизмов, имеющих спирально извитую форму. Название получили за форму клетки (от лат. spira - изгиб). Спирохеты относятся к пор. Spirochaetales, включающему 8 родов: Spirochaetae, Serpulina, Cristispira, Treponema, Brachyspira, Borrelia, Leptospira, Leptonema. Клетки спирохет очень тонкие (0,1-0,6 мкм) и длинные (5-50 мкм) и разделяются на три основные структуры: протоплазменный цилиндр (тело клетки), аксиальную (опорную) нить - аксостиль и трехслойную наружную оболочку. Аксиальная нить состоит из отдельных фибрилл, число которых у разных видов различно: у трепонем и лептоспир - 4, у кристиспир больше 100, у боррелий - до 18. Каждая из фибрилл закрепляется в протоплазменном цилиндре. Химический состав аксиальной нити аналогичен составу жгутиков эубактерий. В структуре клетки спирохет различают клеточную стенку (в отличие от клеточных стенок эубактерий она эластична и не обладает ригидностью), цитоплазматическую мембрану, нуклеоид и мезосомы. Эндоспор, капсул и жгутиков спирохеты не образуют. Размножаются поперечным делением клетки.
Извитая форма клеток спирохет поддерживается натянутой аксиальной нитью, которая у лептоспир и трепонем находится между наружной оболочкой и протоплазменным цилиндром, у кристоспир она винтообразно обвивает клетку в виде тонкой каймы, образуя килевидные выступы - кристы. Если аксиальная нить разрывается, то клетка выпрямляется и принимает форму тонкой длинной нити. Движение спирохет осуществляется за счет сокращения фибрилл аксиальной нити, форма движения винтообразная. Среди спирохет есть сапрофитные формы (род Spirochaetae), обитающие в иловых застойных водах при пониженном содержании кислорода, и патогенные для человека и животных. Так, спирохета Borrelia recurrentis - возбудитель возвратного тифа, Treponema pallidum (бледная спирохета) - сифилиса.
Спириллы и другие изогнутые бактерии. Спиральную форму в несколько завитков имеют бактерии родов Spirillum и Azospirillum. Форма клетки обеспечивается наличием ригидной клеточной стенки. На одном или обоих полюсах клетки спириллы имеют пучок жгутиков, обладают большой подвижностью. Источником углерода и энергии служат отдельные аминокислоты и органические кислоты, углеводы почти не используются. Физиологическим свойством спирилл является склонность к микроаэрофильности. Несмотря на облигатную аэробность они предпочитают среды с низким содержанием кислорода (в 2-7 раз ниже, чем содержится в воздухе). Представителем является Spirillum volutans, вид с очень крупными клетками - до 60 мкм в длину, которые при доступе воздуха не развиваются в чистой культуре.
Бактерии рода Spirillum - сапрофиты, распространены в загрязненных пресных и морских водах; Azospirillum - являются компонентами микробных сообществ различных типов почв, ризосферы и ризопланы растений. Азоспириллы обладают высокой азотфиксирующей активностью, за что род получил название Azospirillum. Фиксацию азота они осуществляют в широком диапозоне pH (5,4-8,5) и температуры (10-40° С). На питательных средах образуют пигментированные колонии красного, розового или желтого цвета.
Изогнутые или спиральные клетки имеют бактерии рода Campylobacter, патогенного для животных. Факультативные аэробы. Энергию получают за счет дыхания или брожения. Среди вибрионов есть патогенные виды - Vibrio cholerae - возбудитель азиатской холеры. Бактерии других родов, как и большинство вибрионов, сапрофиты - обитатели почвы, пресных и морских вод.
Род Bdellovibrio представляют бактерии слегка изогнутой формы, размером 0,3-0,45 мкм в диаметре и 0,8-1,2 мкм в длину. Бактерии снабжены одним полярно расположенным, одетым чехлом жгутиком (рис. 8.1), являются облигатными паразитами других, в основном грамотрицательных, бактерий. Типичным представителем служит Bdellovibrio bacteriovorus, что в переводе с латинского означает пиявка, пожирающая бактерий (bdello - пиявка, vorus - пожирающий).
Бделловибрион обладает необычным циклом развития, включающим множественное
 деление. При столкновении с клеткой-жертвой бдеяловибрион прикрепляется своим безжгути- ковым концом к клеточной стенке жертвы, начинает быстро вращаться вокруг своей оси (скорость вращения около 100 об/с) и в результате пробуравливает клеточную стенку. При этом клетка-жертва округляется, а паразит через отверстие проникает в периплазматическое пространство, где происходит его развитие. Жгутик в процессе проникновения теряется. Одновременно в клетку-жертву может внедряться несколько бделловибрионов. При внутриклеточном развитии питание паразита осуществляется за счет веществ, поступающих из протопласта клетки-жертвы. Растущая клетка паразита удлиняется, приобретает форму длинной нити. Затем происходит множественное деление нити: она сегментируется на множество вибрионоподобных клеток со жгутиком каждая. Число таких клеток может достигать 20- 50 в зависимости от размеров клетки-жертвы. Цикл внутриклеточного развития длится около 4 ч. За это время клетка разрушается и паразит освобождается. Бделловибрионы обладают высокой протеолитической активностью. В качестве источников углерода и энергии используют пептиды и аминокислоты, которые расщепляются в процессе дыхания через цикл три карбоновых кислот. К потреблению углеводов они не способны. Жирные кислоты клетки-жертвы могут прямо включать в липиды, а также превращать в нужные для себя соединения. Кроме типового вида В. bacteriovorus известно еще два вида - В. stolpii и В. starri, названные в честь их первооткрывателей Н. Stolp, М. Starr (1962). Бделловибрионы широко распространены в почвах, сточных водах, пресных и морских водах.
деление. При столкновении с клеткой-жертвой бдеяловибрион прикрепляется своим безжгути- ковым концом к клеточной стенке жертвы, начинает быстро вращаться вокруг своей оси (скорость вращения около 100 об/с) и в результате пробуравливает клеточную стенку. При этом клетка-жертва округляется, а паразит через отверстие проникает в периплазматическое пространство, где происходит его развитие. Жгутик в процессе проникновения теряется. Одновременно в клетку-жертву может внедряться несколько бделловибрионов. При внутриклеточном развитии питание паразита осуществляется за счет веществ, поступающих из протопласта клетки-жертвы. Растущая клетка паразита удлиняется, приобретает форму длинной нити. Затем происходит множественное деление нити: она сегментируется на множество вибрионоподобных клеток со жгутиком каждая. Число таких клеток может достигать 20- 50 в зависимости от размеров клетки-жертвы. Цикл внутриклеточного развития длится около 4 ч. За это время клетка разрушается и паразит освобождается. Бделловибрионы обладают высокой протеолитической активностью. В качестве источников углерода и энергии используют пептиды и аминокислоты, которые расщепляются в процессе дыхания через цикл три карбоновых кислот. К потреблению углеводов они не способны. Жирные кислоты клетки-жертвы могут прямо включать в липиды, а также превращать в нужные для себя соединения. Кроме типового вида В. bacteriovorus известно еще два вида - В. stolpii и В. starri, названные в честь их первооткрывателей Н. Stolp, М. Starr (1962). Бделловибрионы широко распространены в почвах, сточных водах, пресных и морских водах.
Миксобактерии н цитофаги слизеобразующие скользящие бактерии, относящиеся соответственно к порядкам Myxobacteriales и Cytophagales.
Это грамотрицательные, палочковидные или веретенообразные бактерии. Клетки часто вытянуты и заострены на концах, длина их колеблется в пределах 0,7-10 мкм.
Клетки миксобактерий не имеют типичной ригидной клеточной стенки. Клеточная стенка у них тонкая, эластичная, поэтому при движении они могут изгибаться, меняя форму тела. Движение миксобактерий осуществляется по типу скольжения и возможно только на твердом субстрате.
Многие миксобактерий образуют плодовые тела, которые могут быть ярко окрашенными в оранжевый или другой цвет. Плодовые тела имеют разную форму и размеры: от
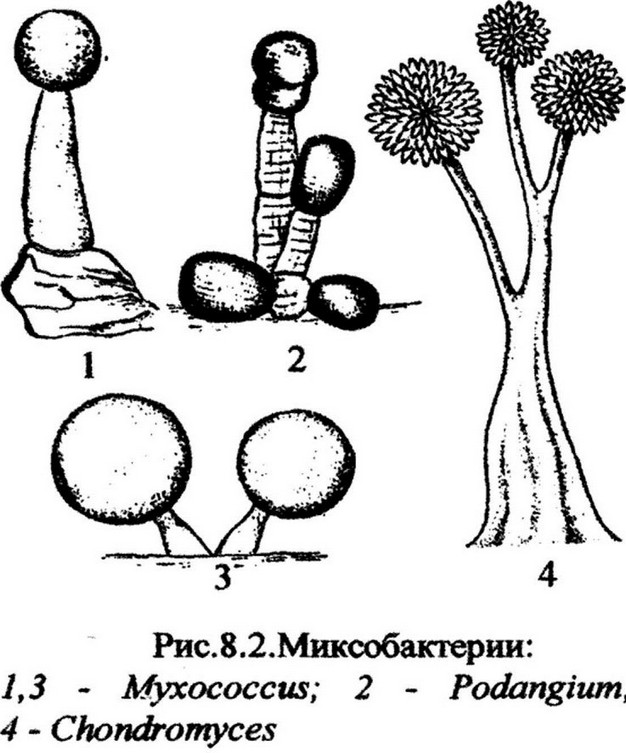 микроскопического бугорка до сложных деревоподобных структур (рис. 8.2). Они представляют собой скопление слизи с погруженными в нее покоящимися укороченными клетками (цисты).
микроскопического бугорка до сложных деревоподобных структур (рис. 8.2). Они представляют собой скопление слизи с погруженными в нее покоящимися укороченными клетками (цисты).
После периода покоя при наличии влаги слизь размягчается, клетки высвобождаются и делятся поперечным делением (перетяжкой).
Цикл развития миксобактерий состоит из двух стадий: стадии роения, или псевдоплазмодия, и стадии
плодоношения, или образования цист. В стадии роения миксобактерий размножаются и выделяют слизистый матрикс (желатинообразную массу), в котором живут и передвигаются клетки. Продолжительность данной стадии 7-8 дней. Для стадии плодоношения характерно образование скоплений клеток в различных точках матрикса. Эти скопления возвышаются над
субстратом, образуя иногда округлые колонии, иногда же слизистый матрикс неравномерно выпячивается, ветвится и дает сложное плодовое тело.
Преобладающее число миксобактерий - сапрофиты. Они распространены в почве, навозе, на разлагающейся древесине, в иле водоемов.
Представители рода Polyangium - активные разрушители целлюлозы. Бактерии рода Myxococcus живут в почве за счет других бактерий, вызывая их лизис выделяемыми литическими ферментами.
Цитофаги в отличие от миксобактерий не образуют плодовых тел. Но главным отличием морфологически сходных этих двух групп бактерий являются нуклеотидный состав ДНК. Содержание ГЦ у цитофаг 30-50 %, у миксобактерий значительно выше - 67-71 %. Группа цитофаг включает три рода: Cytophaga,
Sporocytophaga, Flexibacter. Организмы первых двух родов обитают в почве и вызывают активное расщепление целлюлозы. Некоторые виды рода Cytophaga встречаются как в почве, так и в морской воде. Они вызывают гидролиз хитина и агара.
Представители рода Flexibacter распространены в почве, пресной и морской воде. Среди них есть виды, патогенные для рыб. Так, F. columnaris часто является причиной массовой гибели рыб в рыборазводных прудах.
Энтеробактерии (от греч. entero - кишечник) - своим названием обязаны тому, что большинство видов их является постоянным обитателем кишечного тракта позвоночных. Клетки палочковидной формы, подвижные и неподвижные. Движение осуществляют с помощью полярно или перитрихально расположенных жгутиков. Энтеробактериям свойственна факультативная анаэробность. В анаэробных условиях энергию получают путем сбраживания углеводов, в аэробных - путем кислородного дыхания. Причем в качестве субстратов дыхания могут использовать разнообразные органические соединения. Для энтеробактерий характерно осуществление брожения смешанного типа, при котором образуются молочная, янтарная, уксусная и муравьиная кислоты, этиловый спирт (или углекислый газ) и водород.
Экологически среди энтеробактерий можно выделить три группы: бактерии нормальной микрофлоры млекопитающих и возбудителя кишечных инфекций (роды Escherichia, Salmonella, Shigella), эпифитные и фитопатогенные бактерии (род Erwinia), обитатели воды и почвы (роды Enterobacter, Proteus, Serratia). Все они составляют кишечную группу бактерий.
Род Escherichia - типичный представитель Е. coli (кишечная палочка), входит в состав нормальной микрофлоры кишечного тракта человека и позвоночных животных. Вызыввает заболевания б
исключительных случаях - при ослаблении защитных функций
»
организма.
Е. coli - подвижная палочка, размером 1,1-1,5 - 2,0-6,0 мкм. Хорошо растет на простых питательных средах. Сбраживает углеводы, спирты, некоторые кислоты. Ацетат может использовать как единственный источник углерода, но цитрат использовать не может. Отличительным признаком кишечной палочки от других энтеробактерий является способность быстро (в течение 48 ч) сбраживать лактозу с образованием кислоты и газа, а также образовывать индол из триптофана. Е. coli не нуждается в дополнительных факторах роста. На питательных средах она синтезирует все необходимые аминокислоты.
Е. coli широко используется в санитарной бактериологии как индикатор на загрязнение среды фекалием.
В тесном генетическом родстве с родом Escherichia находятся бактерии родов Shigella и Salmonella. Основным отличительным признаком их является патогенность. Бактерии рода Shigella являются возбудителями дизентерии, разные виды сальмонелл (S. typhi, S. paratyphi) - возбудители брюшного тифа; S. typhimurium. попадая в пищевые продукты, является причиной пищевых токсикоинфекций.
Сальмонеллы - подвижные палочки, снабжены перитрихально расположенными жгутиками, шигеллы подвижностью не обладают Большинство штаммов сальмонелл сбраживают углеводы с образованием кислоты и газа, только S. typhi никогда не образует газ. Шигеллы сбраживают углеводы только с образованием кислоты (без газа).
К энтеробактериям, патогенным для человека и животных, относятся бактерии рода Jersinia, ранее входившие в состав рода Pasteurella. По биохимическим свойствам они сходны с шигеллами. Отдельные виды (J. pestis) неподвижны. Род Jersinia включает всего три вида, которые являются возбудителями заболевания грызунов, а
J. pestis может передаваться от грызунов к человеку и вызывать острое инфекционное заболевание - бубонную чуму. Последняя характеризуется высокой смертностью и относится к группе особо опасных инфекций.
Род Erwinia составляют фитопатогенные и сапрофитные эпифитные бактерии, насчитывающие более 60 видов. Это очень гетерогенная группа, представители которой сходны с другими энтеробактериями не только по морфологии и биохимическим свойствам, но проявляют некоторую способность к гибридизации ДНК, т. е. находятся в определенном генетическом родстве. На основании биохимических критериев род Erwinia разделяют на три подгруппы; Е. amylovora, Е. carotovora, Е. herbicola, названия которым даны по типовым видам.
Бактерии, входящие в состав первой подгруппы, используют ограниченный набор сахаров (ни один вид не использует мальтозу, целлобиозу, лактозу), нуждаются в факторах роста, что не присуще другим бактериям этого рода. Так, характерный представитель группы Е. amylovora испытывает большую потребность в никотиновой кислоте, другие виды развиваются на средах, содержащих дрожжевой экстракт. Бактерии этой подгруппы являются возбудителями ожогов и некротических заболеваний плодовых деревьев.
Подгруппа Е. carotovora (типовой вид Е. carotovora) отличается от других тем, что представители ее обладают способностью синтезировать пектолитические ферменты, которые активно разрушают пектины растений, способствуя этим самым распространению бактерий внутри растений. Е. carotovora поражает многие растения, вызывая мягкую гниль корнеплодов.
Бактерии группы Е. herbicola (типовой вид Е. herbicola) не образуют пектолитических ферментов, являются типичными сапрофитами. В основном обитают на поверхности листьев
Некоторые антимикробные вещества получили широкое практическое применение для подавления роста патогенных микробов. Они называются дезинфицирующими веществами, а прием использования их - дезинфекцией. Наибольшее практическое применение находит хлорная известь (0,5-5 %-ные водные растворы), йод (2 %-ный раствор), двухлористая ртуть, или сулема (1:1000), фенол и его производные (1-5 %-ные растворы), этиловый и изопропиловый спирты (70 %-ные растворы).
клетках позвоночных и членистоногих - клещей, блох, вшей. Наиболее изучены два рода: Rickettsia и Coxiella.
Риккетсии были открыты в 1909 г. американским ученым Гонардом Риккетсом при изучении заболевания пятнистой лихорадки Скалистых гор. Годом позже, работая в Мексике, он установил, что сходный микроорганизм вызывает заболевание - сыпной тиф.
Изучение риккетсий было продолжено чешским ученым Станиславом Провачеком, который, так же как и Г. Риккетс, погиб от сыпного тифа, и бразильским ученым Д. Роха-Лима, предложившим объединить этих неизвестных возбудителей в род Rickettsia (в честь первооткрывателя), а возбудителю сыпного тифа дал видовое название по фамилии чешского исследователя.
Возбудитель сыпного тифа Rickettsia prowazekii паразитирует в клетках эпителия кишечника вшей.
Род Coxiella назван именем американского ученого Г. Кокса, открывшего его. Представители этого рода отличаются от предыдущего меньшими размерами и способностью проходить через бактериальные фильтры. Они являются возбудителями гриппоподобного заболевания, поражающего дыхательные пути, известного под названием лихорадки Q (от англ. Query - неясный, неопределенный). Это заболевание распространено по всему земному шару.
Риккетсии родов Bartonella и Anaplasma паразитируют в эритроцитах человека и других позвоночных, являются одной из причин анемии (малокровие).
Наряду с патогенными известны и непатогенные риккетсии, адаптированные к существованию в членистоногих в основном в качестве симбионтов. Например, организмы рода Symbiotes обитают в мицетомах клопов, рода Blattabacterium - в мицетомах брюшного жирового тела тараканов. Риккетсии не культивируются на синтетических питательных средах. Их можно выращивать в желточном мешке куриных эмбрионов и в культуре клеток.
Хламидии представлены одним родом Chlamidia, имеют сферическую форму клетки, размер которой зависит от стадии развития. Размножаются только в цитоплазме клетки хозяина, и так
называемой цитоплазматической вакуоли, которая образуется в результате инвагинации ЦПМ
Инфекционной формой являются мелкие клетки, диаметром 0,2-0,5 мкм, называемые элементарными тельцами. После проникновения в клетку хозяина они увеличиваются в размерах, превращаясь в крупные сферические клетки (0,8-1,5 мкм) - инициальные тельца. Последние делятся дроблением на дочерние клетки. Цикл завершается преобразованием дочерних клеток в элементарные тельца: уменьшаются размеры клеток, уплотняется ядерный материал, формируется трехслойная клеточная стенка. После разрушения клетки хозяина элементарные тельца проникают в новые клетки и цикл развития повторяется. Некоторое время они могут сохранять жизнеспособность вне клетки хозяина, затем инфицировать его. В отличие от риккетсий хламидии не передаются через беспозвоночных, а непосредственно переходят от одного позвоночного хозяина к другому. Существенным отличием хламидий от риккетсий является также неспособность синтезировать макроэргические соединения, в первую очередь АТФ. В связи с этим их назвали «энергетическими паразитами». Хламидии паразитируют в организме различных позвоночных, в том числе и человека, вызывая инфекционное заболевание - трахому (воспаление слизистой оболочки глаз), пситтакоз, или попугайную болезнь (воспаление дыхательных органов, передающееся человеку от больных попугаев).