
На основании структурно-функциональных свойств автономную нервную систему принято делить на симпатическую, парасимпатическую и метасимпатическую части. Из них первые две имеют центральные структуры и периферический нервный аппарат, мета- симпатическая же часть целиком лежит на периферии в стенках внутренних органов.
Дуга автономного рефлекса (рис. 4.22), как и соматическая рефлекторная дуга, состоит из трех звеньев: чувствительного (афферентного, сенсорного), ассоциативного (вставочного) и эфектор- ного. В зависимости от уровня замыкания, т. е. расположения ассоциативного звена, различают местные, или ганглионарные, спинальные, бульбарные и т. д. рефлекторные дуги. Рефлексы, возникающие при раздражении чувствительных волокон, идущих в составе симпатических и парасимпатических нервов, вовлекают в деятельность не только автономную, но и соматическую нервную систему. Чувствительные волокна этой единой (автономной и соматической) афферентной системы являются отростками биполярных
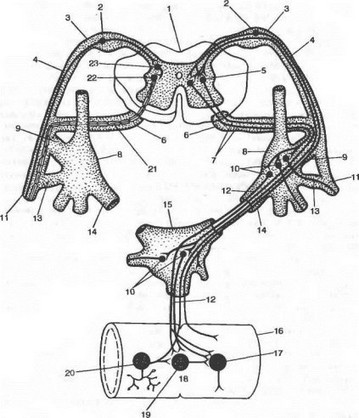
Рис. 4-22. Принципиальная схема дуги автономного (справа) и соматического (слева) рефлексов (по А. Д. Ноздрачеву).
1 — спинной мозг; 2 — тело чувствительного нейрона; 3 — спинальный ганглий; 4 — задний корешок; 5 — вставочный (преганглионарный) нейрон дуги автономного рефлекса; 6 — передний корешок; 7 — прегаглионарное волокно; 8 — околопоэвоночный узел; 9 — белая соединительная ветвь; 10 — двигательный (постганглионарный) нейрон дуги автономного рефлекса; 11 — соматический нерв; 12 — п ос тга игл попарное волокно; 13 — серая соединительная ветвь; 14 — висцеральная ветвь; 15 — пред позвоночный узел (брыжеечного и чревного, или солнечного, сплетения); 16 — кишка; 17 — эффекторный нейрон функционального модуля метасимпатической нервной системы; 18 — функциональный модуль метаем м пяти чес кой нервной системы; 19 — интернейрон функционального модуля; 20 — чувствительный нейрон функционального модуля; 21 — двигательное соматическое волокно; 22 — двигательный нейрон соматической рефлекторной дуги; 23 — вставочный нейрон соматической рефлекторной дуги.
клеток, лежащих в спинномозговых узлах или их аналогах, таких как яремный, тройничный (гассеров) узлы и др. Такое понимание справедливо для сегментарных и рефлекторных дуг более высокого порядка и не относится к местным периферическим дугам автономного рефлекса.
Наряду с общим для обеих (автономной и соматической) систем звеном существует и собственный афферентный путь автономной нервной системы, называемый особым, или висцеральным. Он создает основу для путей местных рефлексов, осуществляемых независимо, без участия ЦНС. По локализации клеточных тел чувствительных нейронов, по ходу и длине отростков их разделяют на три группы. В первую группу объединены клетки, тела которых локализуются в узлах солнечного и нижнего брыжеечного сплетений. Один из их длинных отростков направляется на периферию, другой в сторону спинного мозга. Клетки второй группы характеризуются тем, что их длинный отросток идет к рабочему органу, короткие распределяются в самом ганглии и синаптически контактируют с вставочным или эффекторным нейронами. Висцеральные чувствительные клетки третьей группы отличаются тем, что их тела и короткие отростки располагаются в интрамуральных узлах, длинные же отростки в составе соответствующих нервов достигают симпатических узлов, где и происходит переключение на ассоциативный и моторный (эфферентный) нейрон.
Висцеральная чувствительность обусловлена активностью пяти отдельных типов интероцепторов: механо-, хемо-, термо-, осмо- и ноцицепторов, называемых специфическими. Из них наиболее распространенными являются механорецепторы.
Среди механорецепторов внутренних органов известны рецепторы двух типов: быстро- и медленноадаптирующиеся. Быстро- адаптирующиеся механорецепторы характеризуются высоким порогом возбуждения и встречаются в основном в слизистой оболочке и серозном слое висцеральных органов и связаны преимущественно с миелиновыми волокнами. Характерной чертой быстреадаптирующихся рецепторов являются исключительная чувствительность к динамической фазе движения и сокращения. Для медленноадапти- рующихся механорецепторов, наоборот, характерна генерация сигналов в течение длительного периода раздражения или после его окончания. Эти рецепторы имеются во всех внутренних органах и характеризуются низким порогом возбуждения. Такая особенность позволяет им быть спонтанно-активными и направлять в нервные центры разнообразную информацию о сокращении, расслаблении, растяжении, смещении висцеральных органов. Медленноадаптирующиеся рецепторы связаны с тонкими миелинизированными и без- миелиновыми нервными волокнами.
Хеморецепторы активируются при изменении химического состава ткани, например напряжения С02 и 02 в крови. В органах пищеварения выделены специальные кислого- и щелочечувствительные рецепторы, чувствительные к действию только аминокислот или аминокислот и глюкозы.
Тепловые и холодовые терморецепторы также обнаружены по преимуществу в пищеварительном тракте. Осморецепторы, ионорецепторы (например, натриевые) висцеральных органов обнаружены в печени. Частота их разрядов находится в прямой зависимости от осмотического давления жидкости. Существование специфических ноцицепторов пока еще окончательно не установлено, хотя их роль и отводится некоторым свободным нервным окончаниям. Болевые ощущения возникают при чрезмерной стимуляции любого типа — растяжении, сокращении, действии химических стимулов.
Помимо специфических, имеются и рецепторы, воспринимающие раздражение любой модальности, будь то механическое, химическое, термическое, осмотическое. Местом локализации таких полимодаль- ных интероцепторов является, например, слизистая оболочка пищеварительного тракта.
Все рассмотренные виды висцеральной чувствительности передаются в центры по волокнам трех основных нервных путей: блуждающего, чревных (большого, малого, поясничных) и тазового (рис. 4.23). Из них самым мощным коллектором висцеральной чувствительности является блуждающий нерв. Соотношение в нем афферентных и эфферентных волокон составляет 9:1, в то время как в чревном и тазовом нервах 3:1 и 1:1 соответственно.
Помимо местных сетей, афферентные сигналы могут запускать центральные нейронные механизмы ряда систем: сегментарную, межсегментарную, проприоспинальную, надсегментарную. Несмотря на такую сложность многоступенчатой организации, основа механизма взаимодействия на всех ступенях остается одной и той же: это синаптическая конвергенция к центральным клеткам сигналов различной природы (висцеральной и соматической) и разной модальности, оценка их аппаратом суммации постсинаптических потенциалов и на основании оценки результатов — генерация нового сигнала. Различия между механизмами, включающимися на разных уровнях, непринципиальны и заключаются в количественных особенностях конвергенции. Эти механизмы вовлекаются в разной степени в зависимости от интенсивности воздействия и включаются в разных соотношениях.
Для запуска менее сложных сегментарных механизмов оказывается достаточной и менее сложная суммация постсинаптических процессов. Эти механизмы срабатывают при возбуждении небольшого числа афферентов. Включение более сложных систем требует значительной суммации процессов, а стало быть, более интенсивного притока аффе- рентации. Следовательно, для запуска различных нейронных систем основой является мощность поступающего афферентного потока.
Импульсы, интегрированные в общей системе вставочных нейронов, способны вызвать их активность и, как следствие, появление вегетативных, например дыхательных или сердечно-сосудистых, эффектов. Импульсы могут также активировать клетки высших отделов центральной нервной системы, вызывая появление поведенческих реакций и субъективных ощущений.
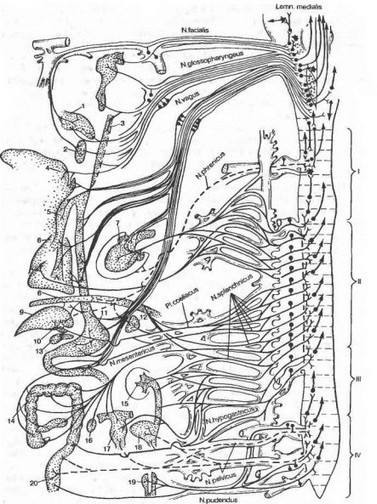
Рис. 4.23. Распределение висцеральных чувствительных волокон в сегментах спинного мозга (по В. Н. Черниговскому). Слева изображены волокна, отходящие от внутренних органов, справа — сегменты спинного мозга и входящие в них волокна.
1 — шейный; II — грудной; III — поясничный; IV — крестцовый отделы спинного мозга; 1 — язычок; 2 — небо; 3 — глотка; 4 — язык; 5 — трахея; б — легкие; 7 — сердце; 8 — диафрагма; 9 — печень; 10 — желчный пузырь; II — желудок; L2 — селезенка; 13 — поджелудочная железа; 14 — тонкая кишка; 15 — почка; 16 — яичник; 17 — матка; 18 — мочевой пузырь; 19 — мочеиспускательный канал; 20 — толстая кишка.
Реакция на афферентный импульс и элементы его переработки на подкорковом уровне является основой для последующих процессов в коре больших полушарий, направленных на регуляцию функций определенной висцеральной системы — пищеварительной, дыхательной и т. д. Эти процессы выражаются в виде вызванных потенциалов — первичного и вторичного ответов: первичные сравниваются с пусковыми, вторичные — с корригирующими влияниями коры большого мозга.
При анализе локализации представительства висцеральных систем в коре большого мозга обнаруживается несоответствие числа зон проекций блуждающего и чревного нервов. Объясняется это тем, что блуждающий нерв по числу сенсорных волокон и особенно по величине иннервируемых областей не имеет себе равных, охватывая большое количество внутренних органов, некоторые из которых подвержены в какой-то мере произвольному контролю.
Представительства функционально близких висцеральных систем находятся и в близко расположенных областях коры. Например, зоны брыжеечных, селезеночных и чревных нервов перекрываются представительством блуждающего нерва, что служит основой для тонкой координации процессов, осуществляемых корой больших полушарий, восстановления функции, надежности работы висцеральных органов.
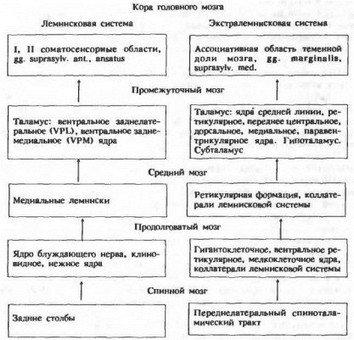
Предложенная В. Н. Черниговским схема проведения висцеральных сигналов в центральной нервной системе дает представление об участии той или иной наиболее важной структуры в этом процессе, хотя и не указывает на степень участия каждой и не отражает всей сложности существующих взаимодействий (схема 4.2). Сигналы, вызывающие ответы в клетках коры большого мозга, после соответствующей обработки передаются в специальные выходы передних отделов поясной извилины, и уже оттуда через гипоталамус нисходящие пути следуют к вставочным (преганглионарным), затем к эффекторным нейронам и далее к исполнительным органам. Таким образом, информация от высших центров по нисходящим путям и от периферических висцеральных и соматических клеток по спинальным дугам поступает к преганглионарным нейронам.
Тело преганглионарного автономного нейрона располагается в сером веществе в одних случаях ствола мозга, в других — спинного мозга. На периферии за пределами спинного мозга нервное волокно вступает в синаптический контакт с эффекторным нейроном. Исключение составляет лишь часть волокон, следующих в составе чревного нерва к надпочечнику. Эти волокна проникают непосредственно в мозговой слой железы, который и выполняет своеобразную функцию постганглионарного звена рефлекторной дуги. Истинное же эффекторное звено дуги автономного рефлекса представляет собой нервную клетку, мигрировавшую из ЦНС.
Преганглионарные волокна различаются по своим функциональным свойствам. Наибольшее их число составляют тонкие, легко возбудимые, с медленным проведением возбуждения единицы. Приближаясь к эффекторным нейронам, преганглионарные волокна те-
Схема 4.2. Проведение висцеральных сигналов по лемиисковой и экстралем- нисковой системам [Черниговский В. Н., 1973|
ряют миелин и разветвляются на тонкие терминалы, образуя на теле и отростках эффекторного нейрона синаптические контакты.
Эффекторных нейронов несравненно больше, чем преганглио- нарных волокон. Например, в верхнем шейном симпатическом ганглии одно преганглионарное симпатическое волокно контактирует более чем с сотней эффекторных нейронов. При этом на одном и том же эффекторном нейроне могут оканчиваться разветвления нескольких преганглионарных волокон. Наличие таких широких конвергентных и дивергентных отношений обеспечивает надежность проведения возбуждения. Эта закономерность касается только симпатической части автономной нервной системы, в двух других ее частях подобная конвергенция практически отсутствует.
В интеграции сигналов в низших центрах вегетативной периферии значительная роль отводится пространственной и временной суммации постсинаптических потенциалов. Роль их состоит в том, что поступающие по пресинаптическим терминалям присущие им относительно слабые сигналы благодаря этим процессам трансфор-
Рис. 4.24. Основные связи автономной нервной системы (no С. П. Семенову).
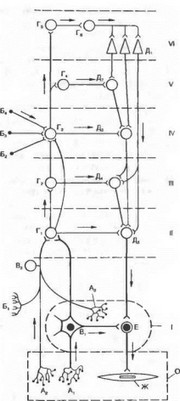 О — висцеральный орган; 1 — автономный (около-, пред позвоночный или интрамуральный) узел; 11 — сегмент спинного мозга или ствола мозга; 111 — надсегментарные центры продолговатого мозга; IV — промежуточный мозг; V — ядра основания мозга; VI — кора больших полушарий головного мозга; А| и Aj — рецепторы внутренних органов; Аз — рецептор автономного нервного узла; Б| — экстероцептор; Бг — Б4 — проводящие пути из органов чувств. В| — чувствительный нейрон автономного нервного узла; Bz — чувствительные нейроны спинномозговых и черепномозговых узлов; Гi — Г4 — ассоциативные ядра разных отделов мозга; Г5 — клетки афферентных зон коры большого мозга; Гб — ассоциативные клетки; Д| — пирамидные клетки коры большого мозга; Дг — ядра основание; Дз — ядра подбугорья; Д4 — клетки ретикулярной формации продолговатого мозга; Дз — ядра автономной нервной системы в стволе мозга и сегментах спинного мозга; Е — эффекториый нейрон дуги автономного рефлекса; Ж — эффекторньк клетки стенки висцерального органа. Стрелки обозначают распространение возбуждения.
О — висцеральный орган; 1 — автономный (около-, пред позвоночный или интрамуральный) узел; 11 — сегмент спинного мозга или ствола мозга; 111 — надсегментарные центры продолговатого мозга; IV — промежуточный мозг; V — ядра основания мозга; VI — кора больших полушарий головного мозга; А| и Aj — рецепторы внутренних органов; Аз — рецептор автономного нервного узла; Б| — экстероцептор; Бг — Б4 — проводящие пути из органов чувств. В| — чувствительный нейрон автономного нервного узла; Bz — чувствительные нейроны спинномозговых и черепномозговых узлов; Гi — Г4 — ассоциативные ядра разных отделов мозга; Г5 — клетки афферентных зон коры большого мозга; Гб — ассоциативные клетки; Д| — пирамидные клетки коры большого мозга; Дг — ядра основание; Дз — ядра подбугорья; Д4 — клетки ретикулярной формации продолговатого мозга; Дз — ядра автономной нервной системы в стволе мозга и сегментах спинного мозга; Е — эффекториый нейрон дуги автономного рефлекса; Ж — эффекторньк клетки стенки висцерального органа. Стрелки обозначают распространение возбуждения.
мируются, превращаясь в сверхпороговые постсинаптические по- тенциалы эфферентного нейрона.
Тело эффекторной клетки дуги автономного рефлекса представляет собой мигрировавшую из спинного мозга клетку, располагающуюся в одном из периферических автономных ганглиев (рис.
4.24). Нейроны этих ганглиев охватывают своим влиянием, как правило, большие территории висцеральных органов. Ганглии могут располагаться либо около позвоночника (превертебральные), либо в сплетениях вблизи внутренних органов (паравертебральные), наконец, в тканях внутренних органов (интрамуральные, интервисцеральные).
Эффекториый нейрон дуги автономного рефлекса по электрическим показателям в покоящемся состоянии мало чем отличается от мотонейрона соматической дуги. Однако кратковременная или одиночная стимуляция преганглионарных волокон вызывает появление в нем сложной последовательности медленных деполяризующих и гиперполяризующих постсинаптических процессов. В этом случае вначале возникает локальный отрицательный потенциал (0-волна), переходящий в положительную П-волну. Последняя сменяется поздней отрицательной П-волной. Каждая из этих фаз отражает межнейронную передачу, при этом 0-волна — возникающий в холинергических синапсах возбуждающий постсинаптический потенциал
(ВПСП). Появление П-волны обусловлено возбуждением особой группы преганглионарных волокон, которые оканчиваются на хромаффинных клетках ганглия. Появление деполяризующихся и ги- перполяризующихся постсинаптических потенциалов опосредуется мускариноподобным действием ацетил холина, в то время как гиперполяризация — специальными вставочными адренергическими клетками, регулируя тем самым возбудимость эффекторных нейронов.
Как правило, эффекторный нейрон может иметь, помимо основного возбуждающего холинергического преганглионарного входа, еще и прямой вход сугубо периферического происхождения, представляющий одно из звеньев местной рефлекторной дуги ганглионарного уровня. Электрофизиологические характеристики эффек- торного нейрона позволяют интегрировать эти сигналы и формировать новый выходной сигнал. Благодаря этим местным дугам в эфферентном нейроне поддерживается необходимый уровень спонтанной активности и при децентрализации ганглия сохраняется его рефлекторная функция. У спонтанно активных эфферентных нейронов фоновые разряды характеризуются низкой частотой. Они могут возникать синхронно пульсовым толчкам, дыхательным и перистальтическим движениям. Паттерн и ритм разрядов совпадают с показателями преганглионарных волокон или активностью волокон местных рефлекторных дуг.