
У позвоночных животных в автономной нервной системе имеется три вида синаптической передачи: электрическая, химическая и смешанная. Органом с типичными электрическими синапсами является цилиарный ганглий птиц, лежащий в глубине глазницы у основания глазного яблока. Передача возбуждения здесь осуществляется практически без задержки в обоих направлениях. К редко встречающимся можно отнести и передачу через смешанные синапсы, в которых одновременно соседствуют структуры электрических и химических синапсов. Этот вид также характерен для цилиарного ганглия птиц. Основным же способом передачи возбуждения в автономной нервной системе является химический. Он осуществляется по определенным закономерностям, среди которых выделяют два принципа. Первый (принцип Дейла) заключается в том, что нейрон со всеми отростками выделяет один медиатор. Как стало теперь известно, наряду с основным в этом нейроне могут при су тс+вовать также другие передатчики и участвующие в их синтезе вещества. Согласно второму принципу, действие каждого медиатора на нейрон или эффектор зависит от природы рецептора постсинаптической мембраны.
В автономной нервной системе насчитывают более десяти видов нервных клеток, которые продуцируют в качестве основных разные медиаторы: ацетилхолин, норадреналин, серотонин и другие биогенные амины, аминокислоты, АТФ. В зависимости от того, какой основной медиатор выделяется окончаниями аксонов автономных нейронов, эти клетки принято называть холинергическими, адренергическими, серотоиинергическими, пуринергическими и т. д. нейронами.
Каждый из медиаторов выполняет передаточную функцию, как правило, в определенных звеньях дуги автономного рефлекса (рис. 4.27). Так, ацетилхолин выделяется в окончаниях всех пре- ганглионарных симпатических и парасимпатических нейронов, а также большинства постганглионарных парасимпатических окончаний. Кроме того, часть постганглионарных симпатических волокон, иннервирующих потовые железы и, по-видимому, вазодилататоры скелетных мышц, также осуществляют передачу с помощью аце- тилхолина. В свою очередь норадреналин является медиатором в постганглионарных симпатических окончаниях (за исключением нервов потовых желез и симпатических вазодилататоров) — сосудов сердца, печени, селезенки.
Медиатор, освобождающийся в пресинапти чески х терминалях под влиянием приходящих нервных импульсов, взаимодействует со спе- цифиеским белком-рецептором постсинаптической мембраны и образует с ним комплексное соединение. Белок, с которым взаимодействует ацетилхолин, носит название холинорецептора, адреналин или норадреналин — адренорецептора и т. д. Местом локализации рецепторов различных медиаторов является не только постсинаптическая мембрана. Обнаружено существование и специальных пресинаптических рецепторов, которые участвуют в механизме обратной связи регуляции медиаторного процесса в синапсе.
Помимо холино-, адрено-, пуринорецепторов, в периферической части автономной нервной системы имеются рецепторы пептидов, дофамина, простагландинов. Все виды рецепторов, вначале обнаруженные в периферической части автономной нервной системы, были найдены затем в пре- и постсинаптических мембранах ядерных структур ЦНС.
Характерной реакцией автономной нервной системы является резкое повышение ее чувствительности к медиаторам после денервации органов. Например, после ваготомии орган обладает повышенной чувствительностью к ацетилхолину, соответственно после симпатэктомии — к норадреналину. Полагают, что в основе этого явления лежит резкое возрастание числа соответствующих рецепторов постсинаптической мембраны, а также снижение содержания или активности ферментов, расщепляющих медиатор (ацетилхолин- эстераза, моноаминоксидаза и др.).
В автономной нервной системе, помимо обычных эффекторыых нейронов, существуют еще специальные клетки, соответствующие
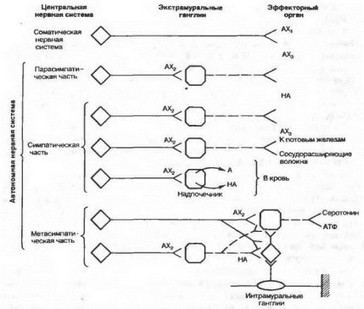
Рис. 4.27. Принципиальная схема химической передачи возбуждения в эфферентном звене рефлекторной дуги соматического и автономного (вегетативного) рефлекса. АХ — аиетилхолии; А — адреналин; НА — норадреналин; АХ| блокируется адом кураре; AXl блокируется ганглиоблокаторами (гексонием); АХз блокируется атропином.
постганглионарным структурам и выполняющие их функцию. Передача возбуждения к ним осуществляется обычным химическим путем, а отвечают они эндокринным способом. Эти клетки получили название трансдукторов. Их аксоны не формируют синаптических контактов с эффекторными органами, а свободно заканчиваются вокруг сосудов, с которыми образуют так называемые гемальные органы. К трансдукторам относят следующие клетки: 1) хромаффинные клетки мозгового слоя надпочечников, которые на холинергический передатчик преганглионарного симпатического окончания отвечают выделением адреналина и норадреналина; 2) юкста- гломерулярные клетки почки, которые отвечают на адренергический передатчик постганглионарного симпатического волокна выделением в кровяное русло ренина; 3) нейроны гипоталамических супраоп- тического и паравентрикулярного ядер, реагирующие на синаптический приток разной природы выделением вазопрессина и оксито- цина; 4) нейроны ядер гипоталамуса.
Действие основных классических меадиаторов может быть воспроизведено с помощью фармакологических препаратов. Например,
никотин вызывает эффект, подобный эффекту ацетилхолина, при действии на постсинаптическую мембрану постганглионарного нейрона, в то время как сложные эфиры холина и токсин мухомора мускарин — на постсинаптическую мембрану эффекторной клетки висцерального органа. Следовательно, никотин вмешивается в межнейронную передачу в автономном ганглии, мускарин — в нейро- эффекторную передачу в исполнительном органе. На этом основании считают, что имеется соответственно два типа холинорецепторов: никотиновые (Н-холинорецепторы) и мускариновые (М-холиноре- цепторы). В зависимости от чувствительности к различным катехоламинам адренорецепторы делят на a-адренорецепторы и р-адренорецепторы. Их существование установлено посредством фармакологических препаратов, избирательно действующих на определенный вид адренорецепторов.
В ряде висцеральных органов, реагирующих на катехоламины, находятся оба вида адренорецепторов, но результаты их возбуждения бывают, как правило, противоположными (табл. 4.2). Например, в кровеносных сосудах скелетных мышц имеются а- и ^-адренорецепторы. Возбуждение «-адренорецепторов приводит к сужению, а /7-адренорецепторов — к расширению артериол. Оба вида адренорецепторов обнаружены и в стенке кишки, однако реакция органа при возбуждении каждого из видов будет одназначно характеризоваться торможением активности гладких мышечных клеток. В сердце и бронхах нет «-адренорецепторов и медиатор взаимодействует только с /7-адренорецепторами, что сопровождается усилением сердечных сокращений и расширением бронхов. В связи с тем что норадреналин вызывает наибольшее возбуждение /7-адренорецепторов сердечной мышцы и слабую реакцию бронхов, трахеи, сосудов, первые стали называть р i-адренорецепторами, вторые — р 2-адренорецепторами.
При действии на мембрану гладкой мышечной клетки адреналин и норадреналин активируют находящуюся в клеточной мембране аденилатциклазу. При наличии ионов Mg2t этот фермент катализирует образование в клетке цАМФ (циклического 3' ,5' -аденозин- монофосфата) из АТФ. Последний продукт в свою очередь вызывает ряд физиологических эффектов, активируя энергетический обмен, стимулируя сердечную деятельность.
Особенностью адренергического нейрона является то, что он обладает чрезвычайно длинными тонкими аксонами, которые разветвляются в органах и образуют густые сплетения. Общая длина таких аксонных терминалей может достигать 30 см. По ходу тер- миналей имеются многочисленные расширения — варикозы, в которых синтезируется, запасается и выделяется медиатор. С приходом импульса норадреналин одновременно выделяется из многочисленных расширений, действуя сразу на большую площадь гладкомы шечной ткани. Таким образом, деполяризация мышечных клеток сопровождается одновременным сокращением всего органа.
Различные лекарственные средства, оказывающие на эффектор- ный орган действие, аналогичное действию постганглионарного волокна (симпатического, парасимпатического и т. п.), получили на-
Таблица 4.2. Показатели изменения функций различных органов при стимуляции симпатических и парасимпатических нервов
|
Орган или система |
Симпатические нервы цепторы |
и ад реноре- |
Парасимпатические нервы |
|
|
нервы |
рецепторы |
|
|
Пищеварительный тракт: |
Ослабление мотори- |
а, р |
|
|
продольные и циркулярные |
Усиление мото- |
||
|
мышцы |
КИ |
|
рики |
|
сфинктеры |
Сокращение |
а |
Расслабление |
|
Мочевой пузырь: |
|
р |
|
|
детрузор |
Расслабление |
Сокращение |
|
|
внутренний сфинктер |
Сокращение |
а |
— |
|
Бронхиальные мышцы |
Расслабление |
Р |
Сокращение |
|
Внутриглазные гладкие мышцы: |
|
|
|
|
мышца, расширяющая зра- |
Сокращение |
а |
— |
|
чок |
|
|
|
|
сфинктер зрачка |
— |
— |
Сокращение |
|
цилиарная мышца |
Расслабление |
Р |
» |
|
Пиломоторные мышцы |
Сокращение |
а |
— |
|
Половые органы: |
|
|
|
|
семенные пузырьки |
» |
» |
— |
|
семявыносящий проток |
» |
» |
— |
|
матка в зависимости от вида |
Расслабление |
р |
— |
|
гормонального фона Сердце: |
|
|
|
|
ритм |
Ускорение |
» |
Замедление |
|
сила сокращения |
Увеличение |
|
Ослаблени |
|
Кровеносные сосуды: |
|
|
|
|
артерии кожи |
Сужение |
— |
— |
|
» брюшной полости |
» |
— |
— |
|
» скелетных мышц |
» |
— |
— |
|
» коронарные |
Сужение, расширение |
а |
— |
|
сосуды мозга |
Сужение |
» |
Расширение (?) |
|
артерии половых органов |
» |
» |
Расширение |
|
веиы |
* |
» |
— |
|
Экзокринные железы: |
|
|
|
|
слюнные |
Секреция |
|
Секреция |
|
слезные |
— |
|
» |
|
пищеварительные потовые |
Снижение секреции Секреция холинергическая |
» |
» |
|
Метаболизм: |
|
|
|
|
печень |
Гликогенолиз, глюконеогенез |
|
|
|
жировые клетки |
Липолиз |
р |
— |
|
секреция инсулина |
Снижение |
» |
— |
звание миметиков (адрено-, холиномиметики). Наряду с этим имеются и вещества, избирательно блокирующие функцию рецепторов постсинаптической мембраны. Они названы ганглиобло- каторами. Например, аммониевые соединения избирательно выключают Н-холинорецепторы, а атропин и скополамин — М-холи- но рецепторы.
Классические медиаторы выполняют не только функцию передатчиков возбуждения, но обладают и общебиологическим действи
ем. К ацетилхолину наиболее чувствительна сердечно-сосудистая система, он вызывает и усиленную моторику пищеварительного тракта, активируя одновременно деятельность пищеварительных желез, сокращает мускулатуру бронхов и понижает бронхиальную секрецию. Под влиянием норадреналина происходит повышение систолического и диастолического давления без изменения сердечного ритма, усиливаются сердечные сокращения, снижается секреция желудка и кишки, расслабляется гладкая мускулатура кишки и т. д. Более разнообразным диапазоном действий характеризуется адреналин. Посредством одновременной стимуляции ино-, хроно- и дромотрепной функций адреналин повышает сердечный выброс. Адреналин оказывает расширяющее и антиспазматическое действие на мускулатуру бронхов, тормозит моторику пищеварительного тракта, расслабляет стенки органов, но тормозит деятельность сфинктеров, секрецию желез пищеварительного тракта.
В тканях всех видов животных обнаружен серотонин (5-ок- ситриптамин). В мозге он содержится преимущественно в структурах, имеющих отношение к регуляции висцеральных функций, на периферии продуцируется энтерохромаффинными клетками кишки. Серотонин является одним из основных медиаторов метасимпати- ческой части автономной нервной системы, участвующей преимущественно в нейроэффекторной передаче, и выполняет также ме- диаториую функцию в центральных образованиях. Известно три типа серотонинергических рецепторов — Д, М, Т. Рецепторы Д-типа локализованы в основном в гладких мышцах и блокируются диэтила мидом лизергиновой кислоты. Взаимодействие серотонина с этими рецепторами сопровождается мышечным сокращением. Рецепторы М-типа характерны для большинства автономных ганглиев; блокируются морфином. Связываясь с этими рецепторами, передатчик вызывает ганглиостимулирующий эффект. Рецепторы Т-типа, обнаруженные в сердечной и легочной рефлексогенных зонах, блокируются тиопендолом. Действуя на эти рецепторы, серотонин участвует в осуществлении коронарных и легочных хеморефлексов. Серотонин способен оказывать прямое действие на гладкую мускулатуру. В сосудистой системе оно проявляется в виде констрикторных или дилататорных реакций. При прямом действии сокращается мускулатура бронхов, при рефлекторном — изменяются дыхательный ритм и легочная вентиляция. Особенно чувствительна к серотонину пищеварительная система. На введение серотонина она реагирует начальной спастической реакцией, переходящей в ритмические сокращения с повышенным тонусом и завершающейся торможением активности.
Для многих висцеральных органов характерной является пури- нергическая передача, названная так вследствие того, что при стимуляции пресинаптических терминалей выделяются аденозин и инозин — пуриновые продукты распада. Медиатором же в этом случае является А Т Ф. Местом его локализации служат пресинаптические терминалы эффекторных нейронов метасимпатической части автономной нервной системы.
Выделившийся в синаптическую щель АТФ взаимодействует с пуринорецепторами постсинаптической мембраны двух типов. Пу- ринорецепторы первого типа более чувствительны к аденозину, второго — к АТФ. Действие медиатора направлено преимущественно на гладкую мускулатуру и проявляется в виде ее релаксации. В механизме кишечной пропульсии пуринергические нейроны являются главной антагонистической тормозной системой по отношению к возбуждающей холинергической системе. Пуринергические нейроны участвуют в осуществлении нисходящего торможения, в механизме рецептивной релаксин желудка, расслабления пищеводного и анального сфинктеров. Сокращения кишечника, возникающие вслед за пуринергически вызванным расслаблением, обеспечивают соответствующий механизм прохождения пищевого комка.
В числе медиаторов может быть гистамин. Он широко распространен в различных органах и тканях, особенно в пищеварительном тракте, легких, коже. Среди структур автономной нервной системы наибольшее количество гистамина содержится в постганглионарных симпатических волокнах. На основании ответных реакций в некоторых тканях обнаружены и специфические гистамино- вые (Н-рецепторы) рецепторы: Н,- и Нгрецепторы. Классическим действием гистамина является повышение капиллярной проницаемости и сокращение гладкой мускулатуры. В свободном состоянии гистамин снижает кровяное давление, уменьшает частоту сердечных сокращений, стимулирует симпатические ганглии.
На межнейронную передачу возбуждения в ганглиях автономной нервной системы тормозное влияние оказывает ГАМК. Как медиатор она может принимать участие в возникновении пресинап- тического торможения.
Большие концентрации различных пептидов, особенно субстанции Р, в тканях пищеварительного тракта, гипоталамуса, задних корешков спинного мозга, а также эффекты стимуляции последних и другие показатели послужили основанием считать субстанцию Р медиатором чувствительных нервных клеток.
Помимо классических медиаторов и «кандидатов» в медиаторы, в регуляции деятельности исполнительных органов участвует еще большое число биологически активных веществ — местных гормонов. Они регулируют тонус, оказывают корригирующее влияние на деятельность автономной нервной системы, им принадлежит существенная роль в координации нейрогуморальной передачи, в механизмах выделения и действия медиаторов.
В комплексе активных факторов видное место занимают проста гландины, которых много содержится в волокнах блуждающего нерва. Отсюда они выделяются спонтанно либо под влиянием стимуляции. Существует несколько классов простагландинов: Е, G, А, В. Их основное действие — возбуждение гладких мышц, угнетение желудочной секреции, релаксация мускулатуры бронхов. На сердечно-сосудистую систему они оказывают разнонаправленное действие: простагландины класса А и Е вызывают вазодилатацию и гипотензию, класса G — вазоконстрикцию и гипертензию.
- Влияние автономной нервной системы на функции тканей
и органов
Главной функцией автономной нервной системы является регулирование процессов жизнедеятельности органов тела, согласование и приспособление их работы к общим нуждам и потребностям организма в условиях окружающей среды. Выражением этой функции служит регуляция метаболизма, возбудимости и других сторон деятельности органов и самой ЦНС. В этом случае управление работой тканей, органов и систем осуществляется посредством двух типов влияний — пусковых и корригирующих.
Пусковые влияния используются в случае, если работа исполнительного органа не является постоянной, а возникает лишь с приходом к нему импульсов по волокнам автономной нервной системы. Если же орган обладает автоматизмом и его функция осуществляется непрерывно, то автономная нервная система посредством своих влияний может усиливать или ослаблять его деятельность в зависимости от потребности. Это будут корригирующие влияния. Пусковые влияния могут дополняться корригирующими.
Влияние автономной нервной системы на висцеральные функции. Все структуры и системы организма иннервируются волокнами автономной нервной системы. Многие из них имеют двойную, а полые висцеральные органы даже тройную (симпатическую, парасимпатическую и метасимпатическую) иннервацию. Изучение роли каждой из них обычно осуществляют с помощью электрического раздражения, хирургического или фармакологического выключения, химической стимуляции и т. д.
Так, сильное раздражение симпатических волокон вызывает учащение сердечных сокращений, увеличение силы сокращения сердца, расслабление мускулатуры бронхов, снижение моторной активности желудка и кишечника, расслабление желчного пузыря, сокращение сфинктеров и другие эффекты (рис. 4.28). Раздражение блуждающего нерва характеризуется противоположным действием: уменьшается ритм и сила сердечных сокращений, расширяются сосуды языка, слюнных желез, половых органов, суживаются бронхи, активизируется работа желудочных желез, расслабляются сфинктеры мочевого пузыря и сокращается его мускулатура (см. табл. 4.2).
Эти наблюдения послужили основанием для представления о существовании «антагонистических» отношений между симпатической и парасимпатической частями автономной нервной системы. Их взаимоотношение уподоблялось коромыслу весов, в которых подъем на определенный уровень одной чаши сопровождается снижением на такой же уровень другой.
Представлению «уравновешивания» симпатических влияний парасимпатическими противоречит ряд фактов: например, слюноотделение стимулируется раздражением волокон симпатической и парасимпатической природы, так что здесь проявляется согласованная реакция, необходимая для пищеварения; ряд органов и тканей снабжается только либо симпатическими, либо парасимпатическими во-
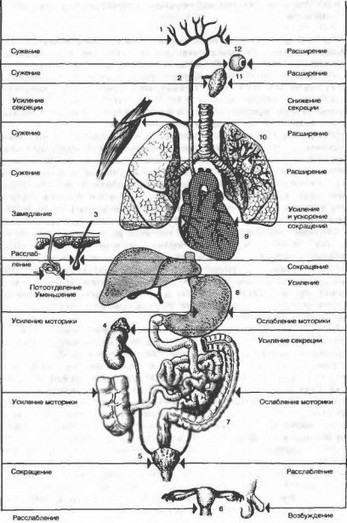
Рис. 4.28. Изменение функций различных органов при раздражении иннервирующих их симпатических (справа) и парасимпатических (слева) волокон.
1 — сосуды головного мозга; 2 — периферические сосуды; 3 — валосянаа мышца; 4 — надпочечник; 5 — мочевой пузырь; б — органы размножение; 7 — кишечник; 8 — желудок; 9 — сердце; 10 — бронхи; 11 — слюнные железы; 12 — зрачок.
локнами. К таким органам относятся многие кровеносные сосуды, селезенка, мозговой слой надпочечника, некоторые экзокринные железы, органы чувств и ЦНС.
Известно, что многие внутренние органы, извлеченные из организма, продолжают выполнять присущие им функции. Например, сохраняется перистальтическая и всасывательная функция кишки и т. д. Такая относительная функциональная независимость объясняется наличием в стенках этих органов метасимпатической части автономной нервной системы, которая обладает собственным нейрогенным ритмом, имеет полный набор необходимых для самостоятельной рефлекторной деятельности звеньев — сенсорного, ассоциативного, эффекторного с соответствующим медиаторным обеспечением. В составе этой системы имеются собственные сенсорные элементы (механо-, хемо-, термо-, осморецепторы), которые посылают в свои внутренние сети информацию о состоянии иннервируемого органа, а также способны передавать сигналы в ЦНС. Сфера иннервации метасимпатической части автономной нервной системы ограничена и охватывает сугубо внутренние органы и то не все. Для этих органов метасимпатическая иннервация является базовой, все звенья ее рефлекторного пути локализуются только в интрамуральных ганглиях. Метасимпатическая часть не имеет своего центрального аппарата и ее эфферентные связи с центральными структурами опосредованы нейронами симпатической и парасимпатической частей автономной нервной системы, образующими синаптические контакты на телах и отростках метасимпатических интернейронов и эффекторных нейронов (см. рис. 4.19).
Мнение о том, что метасимпатическая часть автономной нервной системы является диффузным парасимпатическим ганглием, в котором прямые синаптические контакты между преганглионарными волокнами и ганглионарными клетками являются основой для управления (например, сердечной, желудочной или кишечной функции) , при экспериментальном рассмотрении не подтверждается. Несостоятельно также и представление о метасимпатической части автономной нервной системы как о третьем нейроне в эфферентном звене симпатического пути. Метасимпатическая часть автономной нервной системы — это относительно независимая самостоятельная интегративная система. Ее функцию можно уподобить микропроцессору, расположенному в непосредственной близости от эффекторов (гладкая мышца, всасывающий и экскретирующий эпителий, экзокринные и эндокринные элементы), которые ею контролируются и регулируются.
Невыгодность размещения в ЦНС аппарата, необходимого для постоянного и непрерывного контроля за каждой из висцеральных функций, подтверждается тем, что только метасимпатическая часть автономной нервной системы в кишечнике имеет такое же число клеток (1 ¦ 10е), что и весь спинной мозг, а число метасимпатических нейронов, приходящихся на 1 см2 поверхности кишечника, составляет около 20 000. Существование специальных местных метасимпатических механизмов регуляции функций имеет определенный физиологический смысл. Их наличие увеличивает надежность регуляции функций. Эта регуляция может происходить в случае выключения связи с центральными структурами. При этом ЦНС освобождается от избыточной информации.
Основная функциональная роль метасимпатической части автономной нервной системы состоит в осуществлении механизмов, обеспечивающих гомеостаз — относительное динамическое постоянство внутренней среды и устойчивость основных физиологических функций. В отличие от нее симпатическая часть автономной нервной системы рассматривается как система тревоги, мобилизации защитных сил и ресурсов для активного взаимодействия с факторами среды. Задачу восстановления и поддержания этого постоянства, нарушенного в результате возбуждения симпатической части автономной нервной системы, берет на себя метасимпатическая и отчасти парасимпатическая части автономной нервной системы.
Автономные (вегетативные) рефлексы. Переключение висцеральных афферентных сигналов на эфферентные клетки может происходить в периферических образованиях автономной нервной системы: пара-, превертебральных и интрамуральных ганглиях, называемых низшими рефлекторными центрами, а также на спинальном уровне. В спинальных структурах имеется специальный интернейронный аппарат, осуществляющий первичную обработку сенсорных сигналов. Этот аппарат соединен с клетками боковых рогов спинного мозга и может согласовывать афферентные сигналы, поступающие одновременно из различных рецептивных зон при раздражении интеро- и экстероцепторов. Интеграция висцеральных и соматических сигналов не ограничивается сегментарным спинномозговым уровнем и в определенных условиях может осуществляться уровнями более высокого порядка (см. рис. 4.17). Их координация осуществляется в центрах, расположенных в ретикулярной формации ствола мозга, мозжечке, гипоталамусе, лимбических образованиях и в коре большого мозга.
Процессы в автономной и соматической нервной системе тесно связаны, хотя в ответ на раздражение висцеральных афферентных волокон автономная и соматическая системы вовлекаются в разной степени. Рефлексы в этом случае разделяются на висцеро-висце- ральные, висцеросоматические и висцеросенсорные. Уместно назвать соматовисцеральный рефлекс, а также отметить, что в клинической практике существенное значение отводится еще висцеродермальным и дермовисцеральным рефлексам.
Висцеро-висцеральный рефлекс включает пути, в которых возбуждение возникает и заканчивается во внутренних органах. В этом случае рефлекторные дуги могут быть разного уровня. Одни замыкаются в интрамуральных ганглиях и обеспечиваются метасимпатической иннервацией, другие — в пара- и превертебральных симпатических узлах, наконец, третьи имеют спинальный и более высокий уровень замыкания.
При висцеро-висцеральном рефлексе внутренний орган может отвечать двояко: либо торможением, либо усилением функций.
К числу таких рефлексов относится классический рефлекс Гольца: механическое раздражение брыжейки вызывает замедление частоты сердечных сокращений. Другим примером служит раздражение рецепторов пищеварительного тракта, сопровождающееся ослаблением тонуса мышц, суживающих зрачок. Раздражение каротидной или аортальной рефлексогенных зон влечет за собой изменение интенсивности дыхания, уровня кровяного давления, частоты сердечных сокращений.
Разновидностью висцеро-висцерального является аксон-рефлекс. Это понятие охватывает рефлекторные процессы, осуществляющиеся по разветвлениям аксона без участия тела нервной клетки. Возбуждение возникает в одной ветви аксона, затем переходит на другую и по ней направляется к исполнительному органу, вызывая соответствующую реакцию. Есть и другое объяснение возникновению аксон-рефлекса. Экспериментально доказано, что при возбуждении непосредственно рецепторов из рецепторных мембран выделяются биологически активные вещества типа АТФ и разнообразных пептидов, обладающих вазодилататорным действием, которые вызывают соответствующий эффект.
Понятие аксон-рефлекса используется довольно широко. Им, например, объясняют механизм возникновения сосудистой реакции при раздражении кожных болевых рецепторов. Аксон-рефлекс удается воспроизвести даже после удаления спинного мозга, а также дегенерации симпатических волокон, иннервирующих сосудистую стенку исследуемой области.
Висцеросоматический рефлекс также возникает при раздражении внутренних органов и в дополнение к висцеральным вызывает появление соматических реакций. Они выражаются, например, в изменении текущей активности, сокращении или расслаблении скелетных мышц. Примером такой реакции может служить торможение общей двигательной активности организма при раздражении чувствительных окончаний синокаротидной зоны, а также сокращение мышц брюшной стенки или подергивание конечностей при раздражении рецепторов пищеварительного тракта.
Висцеросенсорный рефлекс осуществляется по тем же путям, что и висцеросоматический, но для его вызова необходимо продолжительное и сильное воздействие. Реакция возникает не только во внутренних органах, соматической мышечной системе, но в дополнение к этому изменяется и соматическая чувствительность. Зона повышенного восприятия обычно ограничивается участком кожи, иннервируемым сегментом, к которому поступают импульсы от раздражаемого висцерального органа. Механизм этого явления основан на том, что висцеральные и кожные чувствительные волокна конвергируют на одних и тех же нейронах спинно-таламического пути, в промежуточных структурах происходит потеря специфичности информации, в результате чего ядерные структуры центральной нервной системы и кора большого мозга связывают возникающее возбуждение с раздражением определенной области кожной поверхности.
Среди рефлексов этого типа особое значение придается висце- родермальному рефлексу, при котором раздражение внутренних органов сопровождается изменением потоотделения, электрического сопротивления (электропроводимости) кожи, изменением кожной чувствительности. Вследствие сегментарной организации автономной и соматической иннервации на ограниченных участках поверхности тела, топография которых различна в зависимости от того, какой орган раздражается, при заболевании внутренних органов возникает повышение тактильной и болевой чувствительности определенных областей кожи. Эти боли названы отраженными, а области их проявления — зонами Захарьина—Геда.
Существует и соматовисцеральный рефлекс, разновидностью которого является дермовисцеральный рефлекс. Он выражается тем, что при раздражении некоторых областей поверхности тела возникают сосудистые реакции и изменения функций определенных висцеральных органов. Это явление послужило основанием для возникновения целого направления клинической медицины — рефлексотерапии.
Адаптационно-трофическая функция симпатической части автономной нервной системы. Л. А. Орбели и сотр. провели исследование функционального значения симпатической иннервации для скелетных мышц, что позволило ему сформулировать учение об адаптационно-трофическом влиянии симпатической части автономной нервной системы. В этом влиянии было выделено два неразрывно связанных компонента: влияния адаптационные и влияния трофические, лежащие в основе адаптационных.
Под адаптационными понимаются влияния симпатической части автономной нервной системы, в результате которых происходит приспособление органов к выполнению тех или иных функциональных нагрузок. Сдвиги наступают благодаря тому, что симпатические влияния оказывают на органы трофическое действие, которое выражается в изменении скорости протекания метаболических процессов.
В 20-х годах А. Г. Гинецинский, изучая влияние симпатических волокон на скелетную мышцу лягушки, обнаружил, что утомленная до полной неспособности сокращаться мышца начинает отвечать на стимуляцию моторных нервов после раздражения ее симпатических волокон вначале слабыми, а потом все более сильными сокращениями (рис. 4.29). Оказалось, что при стимуляции симпатических волокон мышца приобретала способность к развитию более сильного напряжения и более длительного его поддержания даже в условиях тетанического возбуждения. В мышце в этот момент происходят укорочение хронаксии, облегчение перехода возбуждения с нерва на мышцу, повышение чувствительности к ацетил холину, изменение упруговязких свойств и электрической проводимости, повышение потребления кислорода. В миокарде под влиянием раздражения симпатических волокон возникают изменения в потреблении кислорода, содержания гликогена, креатинфосфата, АТФ, актомиозина, РНК, ДНК, фосфолипидов, гуанин-, аденин-, урацилнуклеотидов в активности ряда ферментов.

Рис. 4.29. Влияние раздражения симпатических волокон на кривую мышечного утомления изолированной икроножной мышцы лягушки (по Орбели—Гинецинскому). Сокращения утомляемой мышцы вызываются ритмическим (30 имп/минgt; раздражением соматических двигательных волокон. Моменты раздражения симпатического нерва отмечены поднятием сигнальной линии.
Эти влияния распространяются не только на мышечную деятельность, но относятся к работе рецепторов, синапсов, различных отделов ЦНС, эндокринных желез, к протеканию безусловных спинномозговых, вазомоторных и дыхательных рефлексов, а также условнорефлекторной деятельности. Эффекты адаптационно-трофического влияния, полученные сначала при раздражении симпатических волокон, полностью воспроизводятся раздражением гипоталамич