
Высшим отделом ЦНС является кора большого мозга (кора больших полушарий). Она обеспечивает совершенную организацию поведения животных на основе врожденных и приобретенных в онтогенезе функций.
Кора большого мозга имеет следующие морфофункциональные особенности:
- многослойность расположения нейронов;
- модульный принцип организации;
- соматотопическая локализация рецептирующих систем;
- экранность, т. е. распределение внешней рецепции на плоскости нейронального поля коркового конца анализатора;
- зависимость уровня активности от влияния подкорковых структур и ретикулярной формации;
- наличие представительства всех функций нижележащих структур ЦНС;
- цитоархитектоническое распределение на поля;
- наличие в специфических проекционных сенсорных и моторной системах вторичных и третичных полей с ассоциативными функциями;
- наличие специализированных ассоциативных областей;
- динамическая локализация функций, выражающаяся в возможности компенсаций функций утраченных структур;
- перекрытие в коре большого мозга зон соседних периферических рецептивных полей;
- возможность длительного сохранения следов раздражения;
- реципрокная функциональная взаимосвязь возбудительных и тормозных состояний;
- способность к иррадиации возбуждения и торможения;
- наличие специфической электрической активности.
Глубокие борозды делят каждое полушарие большого мозга на
лобную, височную, теменную, затылочную доли и островок. Островок расположен в глубине сильвиевой борозды и закрыт сверху частями лобной и теменной долей мозга.
Кора большого мозга делится на древнюю (archicortex), старую (paleocortex) и новую (neocortex). Древняя кора наряду с другими функциями имеет отношение к обонянию и обеспечению взаимодействия систем мозга. Старая кора включает поясную извилину, гиппокамп. У новой коры наибольшее развитие величины, дифференциации функций отмечается у человека. Толщина новой коры колеблется от 1,5 до 4,5 мм и максимальна в передней центральной извилине.
Функции отдельных зон новой коры определяются особенностями ее структурно-функциональной организации, связями с другими структурами мозга, участием в восприятии, хранении и воспроизведении информации при организации и реализации поведения, регуляции функций сенсорных систем, внутренних органов.
Особенности структурно-функциональной организации коры большого мозга обусловлены тем, что в эволюции происходила кортикализация функций, т. е. передача коре большого мозга функций нижележащих структур мозга. Однако эта передача не означает, что кора берет на себя выполнение функций других структур. Ее роль сводится к коррекции возможных нарушений функций взаимодействующих с ней систем, более совершенного, с учетом индивидуального опыта, анализа сигналов и организации оптимальной реакции на эти сигналы, формирование в своих и в других заинтересованных структурах мозга памятных следов о сигнале, его характеристиках, значении и характере реакции на него. В дальнейшем, по мере автоматизации реакция начинает выполняться подкорковыми структурами.
Общая площадь коры большого мозга человека около 2200 см2, число нейронов коры превышает 10 млрд. В составе коры имеются пирамидные, звездчатые, веретенообразные нейроны.
Пирамидные нейроны имеют разную величину, их дендриты несут большое количество шипиков; аксон пирамидного нейрона, как правило, идет через белое вещество в другие зоны коры или в структуры ЦНС.
Звездчатые клетки имеют короткие хорошо ветвящиеся дендриты и короткий аскон, обеспечивающий связи нейронов в пределах самой коры большого мозга.
Веретенообразные нейроны обеспечивают вертикальные или горизонтальные взаимосвязи нейронов разных слоев коры.
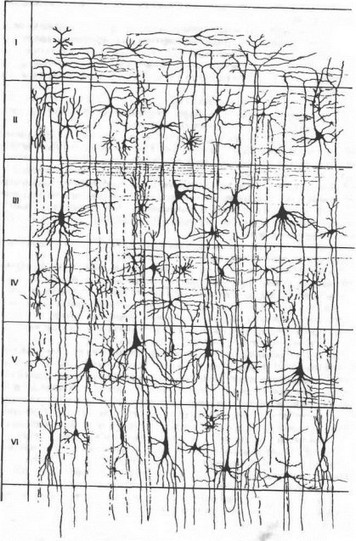
Рис. 4.13. Шестислойное строение коры большого мол га. Объяснение в тексте.
Кора большого мозга имеет преимущественно шестислойное строение (рис. 4.13).
Слой I — верхний молекулярный, представлен в основном ветвлениями восходящих дендритов пирамидных нейронов, среди которых расположены редкие горизонтальные клетки и клетки-зерна, сюда же приходят волокна неспецифических ядер таламуса, регулирующие через дендриты этого слоя уровень возбудимости коры большого мозга.
Слой II — наружный зернистый, состоит из звездчатых клеток, определяющих длительность циркулирования возбуждения в коре большого мозга, т. е. имеющих отношение к памяти.
Слой III — наружный пирамидный, формируется из пирамидных клеток малой величины и вместе со II слоем обеспечивают коркокорковые связи различных извилин мозга.
Слой IV — внутренний зернистый, содержит преимущественно звездчатые клетки. Здесь заканчиваются специфические таламокор- тикальные пути, т. е. пути, начинающиеся от рецепторов анализаторов.
Слой V — внутренний пирамидный, слой крупных пирамид, которые являются выходными нейронами, аксоны их идут в ствол мозга и спинной мозг.
Слой VI — слой полиморфных клеток, большинство нейронов этого слоя образуют кортико-таламические пути.
Клеточный состав коры по разнообразию морфологии, функции, формам связи не имеет себе равных в других отделах ЦНС. Нейронный состав, распределение нейронов по слоям в разных областях коры различны, что позволило выделить в мозге человека 53 ци- тоархитектонических поля. Разделение коры большого мозга на цитоархитектонические поля более четко формируется по мере совершенствования ее функции в филогенезе.
У высших млекопитающих в отличие от низших от двигательного 4 поля хорошо дифференцируются вторичные поля 6, 8 и 10, функционально обеспечивающие высокую координацию, точность движений; вокруг зрительного поля 17 — вторичные зрительные поля 18 и 19, участвующие в анализе значения зрительного стимула (организация зрительного внимания, управление движением глаза). Первичные слуховое, соматосенсорное, кожное и другие поля также имеют рядом расположенные вторичные и третичные поля, обеспечивающие ассоциацию функций данного анализатора с функциями других анализаторов. Для всех анализаторов характерен соматотопический принцип организации проекции на кору большого мозга периферических ре- цептирующих систем. Так, в сенсорной области коры второй центральной извилины имеются участки представительства локализации каждой точки кожной поверхности, в двигательной области коры каждая мышца имеет свою топику (свое место), раздражая которую можно получить движение данной мышцы; в слуховой области коры имеется топическая локализация определенных тонов (тонотопическая локализация), повреждение локального участка слуховой области коры приводит к потере слуха на определенный тон.
Точно так же в проекции рецепторов сетчатки глаза на зри-
тельное поле коры 17 имеется топографическое распределение. В случае гибели локальной зоны поля 17 изображение не воспринимается, если оно падает на участок сетчатки, проецирующийся на поврежденную зону коры большого мозга.
Особенностью корковых полей является экранный принцип их функционирования. Этот принцип заключается в том, что рецептор проецирует свой сигнал не на один нейрон коры, а на поле нейронов, которое образуется их коллатералями и связями. В результате сигнал фокусируется не точка в точку, а на множестве разнообразных нейронов, что обеспечивает его полный анализ и возможность передачи в другие заинтересованные структуры. Так одно волокно, приходящее в зрительную область коры, может активировать зону размером 0,1 мм. Это значит, что один аксон распределяет свое действие на более чем 5000 нейронов.
Входные (афферентные) импульсы поступают в кору снизу, поднимаются к звездчатым и пирамидным клеткам III—V слоев коры. От звездчатых клеток IV слоя сигнал идет к пирамидным нейронам III слоя, а отсюда по ассоциативным волокнам — к другим полям, областям коры большого мозга. Звездчатые клетки поля 3 переключают сигналы, идущие в кору, на пирамидные нейроны V слоя, отсюда обработанный сигнал уходит из коры к другим структурам мозга.
В коре входные и выходные элементы вместе со звездчатыми клетками образуют так называемые колонки — функциональные единицы коры, организованные в вертикальном направлении. Доказательством этого служит следующее: если микроэлектрод погружать перпендикулярно в кору, то на своем пути он встречает нейроны, реагирующие на один вид раздражения, если же микроэлектрод вводить горизонтально по коре, то он встречает нейроны, реагирующие на разные виды стимулов.
Диаметр колонки около 500 мкм и определяется она зоной распределения коллатералей восходящего афферентного таламокор- тикального волокна. Соседние колонки имеют взаимосвязи, организующие участки множества колонок в организации той или иной реакции. Возбуждение одной из колонок приводит к торможению соседних.
Каждая колонка может иметь ряд ансамблей, реализующих какую-либо функцию по вероятностно-статистическому принципу. Этот принцип заключается в том, что при повторном раздражении в реакции участвует не вся группа нейронов, а ее часть. Причем каждый раз часть участвующих нейронов может быть разной по составу, т. е. формируется группа активных нейронов (вероятностный принцип), среднестатистически достаточная для обеспечения нужной функции (статистический принцип).
Как уже упоминалось, разные области коры большого мозга имеют разные поля, определяющиеся по характеру и количеству нейронов, толщине слоев и т. д. Наличие структурно различных полей предполагает и разное их функциональное предназначение (рис. 4.14). Действительно, в коре большого мозга выделяют сенсорные, моторные и ассоциативные области.
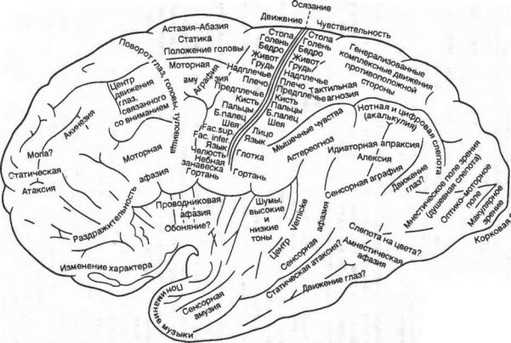
Рис. 4.14. Локализацио функций в коре большого мозга человека (по Экономо и Коскинас).
Г0оота