ГЛАДКАЯ МЫШЕЧНАЯ ТКАНЬ
Основной гистологический элемент гладкомышечной ткани — гладкомышечная клетка (ГМК), способная к гипертрофии и регенерации, а также к синтезу и секреции молекул межклеточного матрикса. ГМК в составе гладких мышц формируют мышечную стенку полых и трубчатых органов, контролируя их моторику и величину просвета. Регуляцию сократительной активности ГМК осуществляют двигательная вегетативная иннервация и множество гуморальных факторов. В ГМК отсутствует поперечная исчерченность, т.к. миофиламенты — тонкие (актиновые) и толстые (миозиновые) нити — не образуют миофибрилл.
А. Миогенез Камбиальные клетки эмбриона и плода (спланхномезодерма, мезенхима, нейроэктодерма) в местах закладки гладкой мускулатуры дифференцируются в миобласты, а затем — в зрелые ГМК, приобретающие вытянутую форму; их сократительные и вспомогательные белки формируют миофиламенты. ГМК в составе гладких мышц находятся в фазе Снеточного цикла и способны к пролиферации.
Б. Гладкомышечная клетка. Морфофункциональная единица гладкой мышечной ткани — ГМК. Заострёнными концами ГМК вклиниваются между соседними клетками и образуют мышечные пучки, в свою очередь формирующие слои гладкой мускулатуры. В волокнистой соединительной ткани между миоцитами и мышечными пучками проходят нервы, кровеносные и лимфатические сосуды. Встречаются и единичные ГМК, например, в субэндотелиальном слое сосудов.
- Общая характеристика. Их форма — вытянутая веретеновидная, часто отростчатая (рис. 7-13). Длина ГМК от 20 мкм до I мм (например, ГМК матки при беременности). Овальное ядро локализовано центрально. В саркоплазме у полюсов ядра расположены хорошо выраженный комплекс Гольджи, многочисленные митохондрии, свободные рибосомы, саркоплазматический ретикулум. Миофиламенты ориентированы вдоль продольной оси клетки. Базальная мембрана, окружающая ГМК, содержит протеогликаны, коллаген типа III и V. Компоненты базальной мембраны и эластин
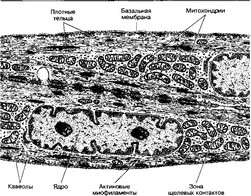
Рис. 7-13. Гладкомышечные клетки. Центральное положение в ГМК занимает крупное ядро. У полюсов ядра находятся митохондрии, эндоплазматический ретикулум и комплекс Гольджи. Актино- вые миофиламенты, ориентированные вдоль продольной оси клетки, прикреплены к плотным тельцам. Миоциты формируют между собой щелевые контакты [из Lentz TL, 1971]
межклеточного вещества гладких мышц синтезируются как самими ГМК, так и фибробластами соединительной ткани.
- Сократительный аппарат. Стабильные актиновые нити ориентированы преимущественно по продольной оси ГМК и прикрепляются к плотным тельцам. Сборку толстых (миозиновых) нитей и взаимодействие актиновых и миозиновых нитей активируют ионы кальция, поступающие из депо Ca2+. Непременные компоненты сократительного аппарата — кальмодулин (Са2+-связывающий белок), киназа и фосфатаза лёгкой цепи гладкомышечного миозина.
- Депо Ca2+ — совокупность длинных узких трубочек (саркоплазматический ретикулум) и находящихся под сарколеммой множества мелких пузырьков (кавеолы). Ca2+- АТФаза постоянно откачивает Ca2+ из цитоплазмы ГМК в элементы саркоплазматического ретикулума. Через Са2+-каналы кальциевых депо ионы Ca2+ поступают в цитоплазму ГМК. Активация Са2+-каналов происходит при изменении мембранного потенциала и при помощи рецепторов рианодина и инозитолтрифосфата.
- Плотные тельца (рис. 7-14). В саркоплазме и на внутренней стороне плазмолеммы находятся плотные тельца — аналог Z-линий поперечнополосатой мышечной ткани. Плотные тельца содержат -актинин и служат для прикрепления тонких (актиновых) нитей.
- Щелевые контакты в мышечных пучках связывают соседние ГМК. Эти нексусы необходимы для проведения возбуждения (ионный ток), запускающего сокращение ГМК.
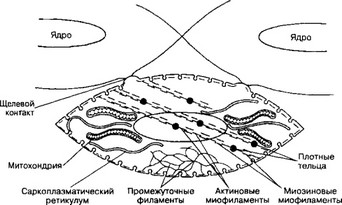
Рис. 7-14. Сократительный аппарат гладкомышечной клетки. Плотные тельца содержат а-актинин, это аналоги Z-линий поперечнополосатой мышцы. В саркоплазме они связаны сетью промежуточных филаментов, в местах их прикрепления к плазматической мембране присутствует винкулин. Актиновые нити прикреплены к плотным тельцам, миозиновые миофиламенты формируются при сокращении [из WestJB, 1990]
В. Механизм сокращения ГМК. В ГМК, как и в других мышечных тканях, работает актомиозиновый хемомеханический преобразователь, но АТФазная активность миозина в гладкомышечной ткани приблизительно на порядок ниже активности АТФазы миозина поперечнополосатой мышцы. Медленное образование и разрушение актин-мио- зиновых мостиков требуют меньшего количества АТФ. Отсюда, а также из факта лабильности миозиновых нитей (их постоянная сборка и разборка при сокращении и расслаблении соответственно) вытекает важное обстоятельство — в ГМК медленно развивается и длительно поддерживается сокращение. При поступлении сигнала к ГМК (через рецепторы плазмолеммы и щелевые контакты) сокращение ГМК запускают ионы кальция, поступающие из кальциевых депо. Рецептор Ca2+ — кальмодулин. Таким образом, увеличение содержания Ca2+ в миоплазме — ключевое событие для сокращения ГМК.
- Регуляция Ca2+ в миоплазме ГМК — процесс, начинающийся с изменения мембранного потенциала и/или связывания рецепторов плазмолеммы с их лигандами (регистрация сигнала) и заканчивающийся изменением режима работы Са2+-кана- лов в депо кальция (открытое или закрытое состояние Са2+-каналов).
а. Изменения мембранного потенциала ГМК происходят при передаче возбуждения от клетки к клетке через щелевые контакты, а также при взаимодействии агонистов (нейромедиаторы, гормоны) с их рецепторами. Изменения мембранного потенциала открывают потенциалзависимые Са2+-каналы плазмолеммы, и в цитоплазме ГМК повышается концентрация Ca2+. Этот Ca2+ активирует рецепторы рианодина кальциевых депо.
б. Рецепторы плазмолеммы ГМК многочисленны {см. III В). При взаимодействии агонистов с их рецепторами (например, норадреналин, гистамин) на внутренней поверхности плазмолеммы активируется фосфолипаза С и образуется второй посредник — инозитолтрифосфат. Инозитолтрифосфат активирует рецепторы ино- зитолтрифосфата кальциевых депо,
в. Активация рецепторов рианодина и инозитолтрифосфата в кальциевых депо открывает их Са2+-каналы, и поступающий в миоплазму Ca2+связывается с кальмодулином.
- Сокращение и расслабление ГМК
а. Сокращение. При связывании Ca2+ с кальмодулином (аналог тропонина С поперечнополосатой мышечной ткани) происходит фосфорилирование лёгкой цепи миозина при помощи киназы лёгких цепей — сигнал для сборки миозиновых нитей и их последующего взаимодействия с тонкими нитями. Фосфорилированный (активный) миозин прикрепляется к актину, головки миозина изменяют свою конформацию, и совершается одно гребковое движение, т.е. втягивание актиновых миофиламентов между миозиновыми. В результате гидролиза АТФ разрушаются актин-миозиновые связи, головки миозина восстанавливают свою конформацию и готовы к образованию новых поперечных мостиков. Продолжающаяся стимуляция ГМК поддерживает формирование новых миозиновых миофиламентов и вызывает дальнейшее сокращение клетки. Таким образом, сила и продолжительность сокращения ГМК определяется концентрацией свободного Ca2+, окружающего миофиламенты.
б. Расслабление. При уменьшении содержания Ca2+ в миоплазме (постоянное откачивание Ca2+ в депо кальция) происходит дефосфорилирование лёгкой цепи миозина при помощи фосфатазы лёгких цепей миозина. Дефосфорилированный миозин теряет сродство к актину, что предотвращает образование поперечных мостиков. Расслабление ГМК заканчивается разборкой миозиновых нитей.
Г. Иннервация. Симпатические (адренергические) и отчасти парасимпатические (холинер- гические) нервные волокна иннервируют ГМК. Нейромедиаторы диффундируют из варикозных терминальных расширений нервных волокон в межклеточное пространство. Последующее взаимодействие нейромедиаторов с их рецепторами в плазмолемме вызывает сокращение либо расслабление ГМК. Существенно, что в составе многих гладких мышц, как правило, иннервированы (точнее, находятся рядом с варикозными терми- налями аксонов) далеко не все ГМК. Возбуждение ГМК, не имеющих иннервации, происходит двояко: в меньшей степени — при медленной диффузии нейромедиаторов, в большей степени — посредством щелевых контактов между ГМК.
Д. Гуморальная регуляция. В мембрану ГМК встроены рецепторы ацетилхолина, гистамина, атриопептина, ангиотензина, адренорецепторы и множество других. Агонисты, связываясь со своими рецепторами в мембране ГМК, вызывают сокращение или расслабление ГМК.
- Сокращение ГМК. Агонист (адреналин, норадреналин, ангиотензин, вазопрессин) через свой рецептор активирует G-белок (Gp)1 который в свою очередь активирует фосфолипазу С. Фосфолипаза С катализирует образование инозитолтрифосфата. Инозитолтрифосфат стимулирует высвобождение Ca2+ из кальциевых депо. Повышение концентрации Ca2+ в саркоплазме вызывает сокращение ГМК.
- Расслабление ГМК. Агонист (атриопептин, брадикинин, гистамин, VIP) связывается с рецептором и активирует G-белок (Gs), который в свою очередь активирует аденилат- циклазу. Аденилатциклаза катализирует образование цАМФ. цАМФ усиливает работу кальциевого насоса, закачивающего Ca2+ в депо кальция. В саркоплазме снижается концентрация Ca2+, и ГМК расслабляется.
ГМК разных органов различно реагируют (сокращением либо расслаблением) на одни и те же лиганды. Это обстоятельство объясняется тем, что существуют разные подтипы конкретных рецепторов с характерным распределением в разных органах.
а. Гистамин действует на ГМК через рецепторы двух типов: H1 и H2.
- Бронхиальная астма. Выброшенный из тучных клеток при их дегрануляции гистамин взаимодействует с Н^гистаминовыми рецепторами ГМК стенки бронхов и бронхиол, что приводит к их сокращению и сужению просвета бронхиального дерева — бронхоспазм.
- Анафилактический шок. Выделяющийся в ответ на аллерген из базофилов гистамин активирует рецепторы типа H1 в ГМК артериол, это вызывает их расслабление, что сопровождается резким падением АД (коллапс).
б. Норадреналин, выделяющийся из симпатических нервных волокон, взаимодействует с ГМК через адренорецепторы двух типов: аир.
- Вазоконстрикция. Норадреналин взаимодействует с а-адренорецепторами ГМК стенки
артериол, что приводит к сокращению ГМК, вазоконстрикции и повышению АД.
- Перистальтика кишечника. Адреналин и норадреналин подавляют перистальтику кишечника, вызывая расслабление ГМК через а-адренорецепторы.
Е. Типы миоцитов. В основе классификации ГМК находятся различия в их происхождении, функциональных и биохимических свойствах.
- Висцеральные ГМК происходят из мезенхимных клеток спланхнической мезодермы и присутствуют в стенке полых органов пищеварительной, дыхательной, выделительной и половой систем. Многочисленные щелевые контакты компенсируют сравнительно бедную иннервацию висцеральных ГМК, обеспечивая вовлечение всех ГМК в процесс сокращения. Сокращение ГМК медленное, волнообразное. Промежуточные филаменты образованы десмином и виментином.
- ГМК кровеносных сосудов развиваются из мезенхимы кровяных островков. Сокращение ГМК сосудистой стенки опосредуют иннервация и гуморальные факторы. Промежуточные филаменты содержат как десмин, так и виментин.
- ГМК радужной оболочки имеют нейроэктодермальное происхождение. Они формируют мышцы, расширяющие и суживающие зрачок. Мышцы получают вегетативную иннервацию. Двигательные нервные окончания подходят к каждой ГМК. Мышца, расширяющая зрачок, получает симпатическую иннервацию из пещеристого сплетения, волокна которого транзитом проходят через ресничный ганглий. Мышца, суживающая зрачок, иннервирована постганглионарными парасимпатическими нейронами ресничного ганглия. На этих нейронах оканчиваются преганглионарные парасимпатические волокна, проходящие в составе глазодвигательного нерва.
- По функциональным свойствам различают тонические и фазные ГМК. Агонист в тонических ГМК вызывает постепенную деполяризацию мембраны (ГМК пищеварительного тракта). Фазные ГМК (vas deferens) генерируют потенциал действия и имеют относительно быстрые скоростные характеристики.
Ж. Регенерация. Вероятно, среди зрелых ГМК присутствуют недифференцированные предшественники, способные к пролиферации и дифференцировке в дефинитивные ГМК. Более того, дефинитивные ГМК потенциально способны к пролиферации. Новые ГМК возникают при репаративной и физиологической регенерации. Так, при беременности в миометрии происходит не только гипертрофия ГМК, но и значительное увеличение их общего количества.
Источник: Под ред. Э.Г. Улумбекова, Ю.А. Челышева, «Гистология (введение в патологию)» 1997