
А. Принципы организации: пограничное расположение (см. А I), пространственная геометрия (А 2 и А 7), практическое отсутствие межклеточного вещества (А 3), полярная дифференцировка (А 4), базальная мембрана (А Б), отсутствие кровеносных сосудов (А 6), выраженная способность к регенерации пограничных эпителиев (А 8).
- Пограничное расположение. Эпителии отделяют организм от внешней среды и от вторичных полостей тела.
- Пласт клеток. Образуя непрерывный слой, эпителий отделяет подлежащие ткани от внешней среды и от вторичных полостей тела. Толщина пластов различна. Например, эпидермис кожи имеет толщину до нескольких десятков микрон, тогда как эпителий на поверхности альвеол лёгкого — около 0,2 мкм. Пласт — не единственный тип организации эпителиев (см. I А 7).
- Незначительные межклеточные пространства. В эпителии практически нет межклеточного вещества, клетки плотно примыкают одна к другой и связаны при помощи специализированных межклеточных контактов (I Б 2, глава 4 IV).
- Полярная дифференцировка эпителиальных клеток. Базальная и апикальная части клетки различаются как структурно, так и функционально. Этот признак обязателен для однослойных эпителиев пограничного расположения (на границе внешней и внутренней сред, на поверхности серозных оболочек), а также для эпителиальных клеток, находящихся в тесной связи с кровеносными капиллярами (эндокринные железы, печень).
а. Апикальная часть содержит микроворсинки, стереоцилии, реснички, секреторный материал и участвует в образовании плотных и промежуточных контактов.
(!) Микроворсинки присутствуют в эпителиальных клетках, осуществляющих транспорт из внешней среды (например, всасывание в кишечнике, реабсорбция в канальцах почки). Основная функция микроворсинок — увеличение площади контакта. Характерные черты микроворсинок — наличие систем транспорта и некоторая их подвижность за счёт актиновых микрофиламентов.
(а) Актиновые микрофиламенты расположены на расстоянии 10 нм друг от друга и соединены в единую систему (стержень микроворсинки) при помощи актин-связывающих белков фимбрина и фасцина (рис. 5-1). Актин периферически расположенных микрофиламентов может взаимодействовать с сократительным белком (минимиозин), расположенным под клеточной мембраной. Микрофиламенты микроворсинок соединены с микрофиламентами, ориентированными параллельно апикальной поверхности клетки; они также связаны с клеточной мембраной при помощи белка спектрина.
Виллин. Микроворсинки эпителиальных клеток пищеварительной, мочевыделительной и половой систем содержат актин-связывающий белок виллин. При болезни Дэвидсона (аутосомное рецессивное наследование дефектного гена виллина) возникает атрофия микроворсинок каёмчатых клеток кишечника.
(б) Транспортные белки
- Глюкоза. В эпителиоцитах, транспортирующих глюкозу из апикальной в базальную часть, в плазматическую мембрану апикальной части встроены переносчики глюкозы (см. главу 2 I В I б (I) (a) (i)).
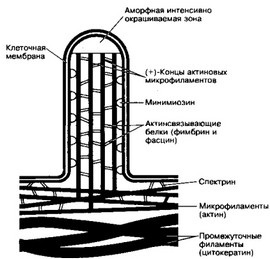
Рис. 5-1. Организация микроворсинки в апикальной части каёмчатой клетки. Около 30 параллельно идущих микрофиламентов образуют стержень микроворсинки. (+)-Концы двух переплетённых нитей F-актина микрофиламентов направлены к вершине микроворсинки. Микрофиламенты заякорены цитоплазматическими концами в терминальной сети. Терминальная сеть — густое сплетение молекул спектрина, сшивающих примембранные микрофиламенты. Непосредственно под терминальной сетью расположено сплетение промежуточных филаментов. Микрофиламенты скрепляют актин- связывающие белки фимбрин и фасцин. Микрофиламенты присоединены к внутренней поверхности плазматической мембраны при помощи минимиозина (из StevensA, Lowe J, 1992]
Oi) Cl' и Na+. В плазматической мембране апикальной части каёмчатых клеток крипт тонкой кишки присутствуют системы транспорта ионов Cl' и Na* из клетки в просвет органа (глава 12 IX Б I а). Нарушение функционирования этих систем вызывает диарею.
- Стерсоцилии (см. главы 8.4 IV Б 3 а, 8.4 IV В I).
- Мерцательные реснички (см. главы 2 III А 6,13 В I).
- Секреторный материал (железистый эпителий желудка, бокаловидная клетка).
- Межклеточные контакты.
б. Базальная часть содержит различные органеллы.
- Митохондрии. Локализация митохондрий преимущественно в базальной части связана с необходимостью АТФ для встроенных в плазмолемму этой части клетки ионных насосов (например, Ыа*,К+-АТФаза).
- Рецепторы гормонов и факторов роста.
- Транспортные системы ионов и аминокислот. Переносчики глюкозы (обеспечивают выход глюкозы из клетки по концентрационному градиенту) отличаются от встроенных в апикальную мембрану.
- Цитоскелет. Полярная дифференцировка проявляется в характере распределения белков, связанных с цитоскелетом. Так, в базальной части преобладают анки- рин и фодрин, локализующиеся совместно с Ыа+,К+-АТФазой.
- Межклеточные контакты. Как правило, это полудесмосомы, связывающие клетки эпителия с базальной мембраной.
в. Липидный состав плазмолеммы апикальной и базальной частей эпителиальных клеток существенно различается. Так, оболочка проникшего в клетку вируса — фрагмент плазмолеммы той части клетки, где вирус проник в клетку. Если вирус проходит в клетку через апикальную часть, то его оболочка содержит преимущественно фосфатидилэтаноламин и фосфатидилсерин — липиды плазмолеммы апикальной части. Если вирус проникает через базальную часть, то его оболочка обогащена другими липидами (фосфатидилхолин, сфингомиелин и фосфатидилинозитол), преобладающими в плазмолемме базальной части.
- Базальная мембрана (пластинка) имеет толщину 20-100 нм, отделяет эпителий от подлежащей соединительной ткани, укрепляет эпителиальный пласт, образуется за счёт эпителия и подлежащей соединительной ткани. Эпителиальные клетки прикреплены к базальной мембране при помощи полудесмосом. Через базальную мембрану осуществляется питание эпителия. Содержит коллаген IV типа (см. главу 6.2 Б 16 (I) (б)), ламинин, энтактин и протеогликаны. У эпителиальных клеток печени нет базальной мембраны.
а. Коллаген IV типа состоит из трёх a-цепей. Две из них (а,- и а -цепи) облигатны для молекулы коллагена IV типа, а одна из цепей (а3, а4, а5 и а6) специфична для эпителиев разной локализации.
Дефектные гены, кодирующие синтез цепей а3, а4, а5 и а6, — причина многих патологических состояний (например, синдром Олпорта, диффузный лейомиоматоз).
б. Ламинин — крупномолекулярный белок базальной мембраны — служит посредником между коллагеном IV типа и эпителиальными клетками. Нарушения структуры ламинина найдены при некоторых заболеваниях — вялая кожа (cutis laxa) и врождённый буллёзный эпидермолиз.
в. Энтактин. Этот гликопротеин связывает коллаген IV типа и ламинин.
- Отсутствие кровеносных сосудов. Некоторые структуры лишены кровеносных сосудов. Типичные примеры: хрящи (глава 6.3 А), средняя (tunica media) и внутренняя (t. intima) оболочки кровеносных сосудов (глава 10 А), пласты эпителия. Метаболизм таких образований (в т.ч. их питание) осуществляется путём диффузии: в хряще — через кровеносные сосуды надхрящницы, в кровеносных сосудах — со стороны наружной оболочки (t. adventitia) и из просвета сосуда.
а. Метаболизм. Питание эпителия, транспорт газов, выведение продуктов метаболизма из эпителия осуществляются путём диффузии веществ через базальную мембрану между эпителием и подлежащими кровеносными сосудами.
б. Причины. Хрящевые клетки вырабатывают факторы, тормозящие ангиогенез (глава 10 А 8). Причина отсутствия кровеносных сосудов в пластах эпителия не очень ясна; возможно механическое препятствие (базальная мембрана и межклеточные контакты между базальными частями эпителиоцитов).
- Эпителиоциты печени лишены базальной мембраны и непосредственно контактируют с кровью, находящейся в пространствах Дйссе. В этом смысле гепатоциты — исключение из общего правила (отсутствие кровеносных сосудов в эпителии).
- Малигнизация. Целостность базальной мембраны и межклеточных контактов в эпителиальных злокачественных опухолях (карциномы) нарушаются, и кровеносные сосуды прорастают в эпителиальную опухолевую ткань.
- Пространственная организация. Эпителиальные клетки организованы в ассоциаты на границе внутренней и внешней среды организма, а также во внутренней среде следующим образом: пласт, тяж, островок, фолликул, трубочка, сеть.
а. Пласт. Эпителиальные клетки, формирующие пласты, всегда имеют пограничное положение (например, эпидермис [часто обозначают как покровный эпителий], эпителии слизистой оболочки кожного и кишечного типа, мезотелии). В силу пограничного положения для клеток однослойного пласта характерна полярная дифференцировка, а многослойные пласты имеют значительные отличия морфологии эпителиальных клеток разных слоёв. Для пластов эпителия характерна выраженная способность к регенерации.
б. Трубочка. Микроскопический вариант пласта, свёрнутого в трубочку (например, потовые железы, канальцы нефрона). Выражена полярная дифференцировка, способность к регенерации определяется генезом эпителия.
в. Островок. Эпителиальные островки всегда погружены во внутреннюю среду организма и, как правило, выполняют эндокринную функцию (например, островки Лангерханса поджелудочной железы). В норме эндокринные клетки островков не регенерируют.
г. Фолликул — имеющие полость островки эпителия. Типичный пример — фолликулы щитовидной железы. Характерна полярная дифференцировка.
д. Тяж. По принципу анастомозирующих тяжей из эпителиальных гепатоцитов организована паренхима печени.
е. Сеть. В вилочковой железе поддерживающий каркас состоит из отростчатых эпителиальных клеток.
- Способность к регенерации выражена у покровных эпителиев и вытекает из их пограничного расположения. Необходимые условия для регенерации — доказанное или предполагаемое наличие стволовых клеток (эпидермис, эпителии слизистой оболочки трубчатых и полостных органов, мезотелии), возможность репликации ДНК с последующим цитокинезом или без него (например, гепатоциты). У погружённых во внутреннюю среду эпителиальных клеток регенераторные возможности существенно меньше вплоть до полной невозможности регенерации (например, р-клетки островков поджелудочной железы). Для ряда эпителиев (например, канальцы нефрона) и клеток эпителиального генеза (например, передняя доля гипофиза) способность к регенерации как будто имеется, хотя её механизмы неясны.