
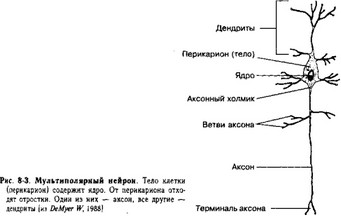
Нейроны (термин предложил Вильгельм фон Вальдейер) — главные клеточные типы нервной ткани. Эти возбудимые клетки осуществляют передачу электрических сигналов (между собой при помощи нейромедиаторов в синапсах) и обеспечивают способность мозга к переработке информации. Существенная часть каждого нейрона — цитоскелет. Перикарион (тело) и отходящие от него отростки (аксон и ветвящиеся дендриты) — стандартные части нейронов (рис. 8-3).
А. Перикарион содержит ядро, комплекс Гольджи, гранулярную эндоплазматическую сеть, митохондрии, лизосомы, элементы цитоскелета.
- Ядро нейрона имеет мелкодисперсный хроматин и ядрышко. В силу относительно большого диаметра ядро в CM выглядит (особенно в крупных нейронах) как оптически пустое. Ядрышко крупное и резко базофильное.
- Комплекс Гольджи хорошо развит, особенно в крупных нейронах. Его особенность — расположение между ядром и местом отхождения аксона, что отражает мощный
транспорт белков, синтезированных в гранулярной эндоплазматической сети перикари- она, в аксон.
- Аксонный холмик — занятая комплексом Гольджи область перикариона, место генерации потенциалов действия.
- Гранулярная эндоплазматическая сеть. В перикарионе и дендритах развита гранулярная эндоплазматическая сеть (глыбки рибосом впервые обнаружил Франц Нисслъ при окраске метиленовым синим, поэтому её в нейронах иногда называют веществом Нйссля [тигроид[††]]).
Тигролиз — распыление тигроидного вещества, отражающее глубокие дистрофические изменения при нарушении целостности нейрона (например, при сдавлении или перерезке аксона).
- Митохондрии многочисленны. Значительные энергетические потребности нервных клеток обеспечивает преимущественно аэробный метаболизм, поэтому нейроны крайне чувствительны к гипоксии.
- Цитоскелет (см. III Д).
- Пигменты. В нейронах (особенно с возрастом) накапливается липофусцин. Нейроны некоторых ядер мозга в норме содержат иные пигменты, поэтому эти образования и получили своё название (substantia nigra, locus coeruleus).
Б. Отростки, отходящие от перикариона, — аксон и дендриты (рис. 8-3 и 8-4). Отростки
нейрона участвуют в образовании синапсов.
- Аксон (нейрит) — длинный отросток, как правило, не ветвящийся по его протяжению, но образующий концевые разветвления, содержащие синаптические пузырьки; проводит пачки импульсов (спайки) от перикариона.
а. Экспрессия нейромодулина (GAP-43) — специфичного для аксона фосфобелка — признак начала дифференцировки нейронов. Сначала образуются короткие отростки, которые потенциально могут стать либо аксоном, либо дендритами. Отросток, накапливающий белок GAP-43, в дальнейшем становится аксоном.
б. Объём аксона может достигать 99% суммарного объёма нейрона.
в. Длина аксона может быть весьма значительной — десятки сантиметров.
- Дендриты — ветвящиеся отростки, заканчивающиеся вблизи от тела нейрона. В плазмолемму встроены постсинаптические рецепторы, дендриты проводят возбуждение к перикариону. Проксимальные области дендритов — продолжение перикариона. Поэтому они содержат рибосомы, компоненты гранулярной и гладкой эндоплазматической сети, элементы комплекса Гольджи.
В. Классификация. Нейроны отличаются по размерам и форме перикариона, числу отростков, их синаптическим связям, характеру ветвления дендритов, электрофизиологическим характеристикам, химии нейромедиаторов, позиции в функциональных сетях и множеству других характеристик. По этой причине классификации нейронов многочисленны.
- Клеточный тип. Эта классификация могла бы быть главенствующей, но находится в стадии разработки.
- Количество отростков
а. Аполяры — отростков нет (условно к ним можно отнести ранние нейробласты).
б. Униполяры — единственный отросток (формально одноотростчатыми нервными клетками можно считать псевдоуниполярные нейроны спинномозговых узлов).
Псевдоуниполяры на самом деле имеют два отростка (центральный и периферический), отходящие от короткого выроста перикариона. В нейроонтогенезе от перикариона отходят два отростка, они сближаются и образуют общий ствол отхождения от перикариона. Периферический отросток иногда называют аксоном, центральный — дендритом, что неверно.
в. Биполяры имеют аксон и ветвящийся дендрит (например, обонятельные рецепторные нервные клетки).
г. Мультиполяры. Число отростков более двух (один аксон, остальные — дендриты). Классический пример — мотонейроны передних рогов спинного мозга.
- Химия нейромедиатора. Критерий классификации — синтез, накопление в синаптических пузырьках и экскреция в синаптическую щель конкретного нейромедиатора. При этом к имени нейромедиатора добавляют ергический. Иногда в качестве критерия применяют тип мембранного рецептора, регистрирующего наличие нейромедиатора (в этом случае добавляют цептивный).
а. Холинергические. Нейромедиатор — ацетилхолин (например, двигательные нейроны передних рогов спинного мозга, иннервирующие скелетные мышечные волокна; парасимпатические нейроны блуждающего нерва, иннервирующие сердце, ГМК и железы желудка).
б. Адренергические. Нейромедиатор — норадреналин (например, постганглионарные нейроны симпатического отдела вегетативной нервной системы, иннервирующие сердце, ГМК сосудов и внутренних органов).
в. Дофаминергические (например, некоторые нервные клетки базальных ядер мозга). Недостаточная секреция дофамина приводит к развитию паркинсонизма.
- Форма перикариона (например, пирамидные и звёэЬштые нейроны коры большого мозга).
- Длина аксона (например, короткоаксонные и длинноаксонные нервные клетки коры больших полушарий). В зависимости от длины аксона различают клетки Гольджи I и II типа. Клетки Гольджи I типа имеют длинные аксоны (например, аксоны пирамидных нейронов коры больших полушарий достигают длины 50-70 см). Клетки Гольджи II типа имеют короткие аксоны. В сером веществе спинного мозга короткие безмиелино- вые аксоны клеток Гольджи II типа могут не выходить за пределы сегмента, проходить в спайках или соединять соседние сегменты. Другим примером клеток Гольджи II типа могут служить вставочные нейроны зернистого слоя коры мозжечка (глава 8.3 IIА 3 б). Вставочные нейроны сетчатки (амакринные клетки [глава 8.4 IА 2 е (3) (б)]) вообще не имеют аксонов.
- Позиция в нейронной цепочке (в т.ч. в дуге рефлекса) позволяет выделять чувствительные (воспринимающие сигнал из внешней или внутренней среды), двигательные (иннервирующие сократительные и секреторные элементы) и находящиеся между ними вставочные (ассоциативные в нейронных сетях) нервные клетки.
- Направление возбуждения к центру — афферентные нервные клетки (в т.ч. чувствительные нейроны разных модальностей, восходящих путей), к периферии — эфферентные нейроны двигательных путей и трактов (например, пирамидной и экстра- пирамидной систем).
- Модальность — характер воспринимаемого и передаваемого сигнала (например, меха- норецепторные, зрительные, обонятельные нейроны и т.д.).
- Отдел нервной системы. Целесообразно выделять нервные клетки вегетативного отдела нервной системы. Нейроны соматического отдела — чувствительные и двигательные, не относящиеся к вегетативным.
- Бддиана т.н. универсальная классификация частей нейрона предложена для сопоставления частей нейрона (перикарион, дендриты, аксон), направления возбуждения и характера электрогенеза в частях нервной клетки (рис. 8-4).
Г. Синапсы (см. также главу 8.2 IIl Б I, рис. 8-20) — специализированные межклеточные
контакты, передающие сигналы от одного нейрона к другому при помощи нейромедиаторов.
Химическая природа нейромедиатора, морфология синапсов и участвующие в формировании
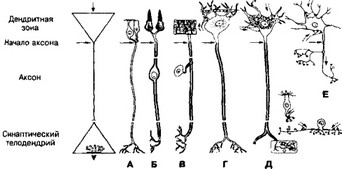
Рис. 8-4. Универсальная классификация частей различных типов нейронов. Дендритная зона — возбуждающий или тормозящий вход. Аксон — проводящая возбуждение (спайки) часть нейрона, его концевые разветвления (выход) участвуют в образовании синапсов с химической или электрической передачей; А-Г — афферентные (чувствительные) нейроны; Д — мотонейрон; E — вставочные (ассоциативные) нейроны [из Bodian D, 1966]
синапса части нейронов в различных отделах нервной системы значительно варьируют. В синапсе выделяют пресинаптическую и постсинаптическую части, разделённые синаптической щелью шириной 20-30 нм. Пресинаптические нейроны синтезируют, хранят и секретируют нейромедиаторы. При изменении мембранного потенциала в терминалях нейромедиатор выделяется в синаптическую щель (экзоцитоз) и связывается со своими рецепторами в постсинаптической мембране, вызывая изменение мембранного потенциала постсинаптического нейрона.
- Классификация
а. Аксодендритические — синапсы между аксоном одного нейрона и дендритами другого нейрона.
б. Аксо-аксональные — синапсы между аксонами разных нейронов.
в. Аксосоматические — синапсы между терминалями аксона одного нейрона и телом другого нейрона.
г. Дендродендритические — синапсы между дендритами нейронов.
- Пресинаптическая часть — специализированная часть терминали отростка нейрона, где расположены синаптические пузырьки и митохондрии. Пресинаптическая мембрана (плазмолемма) содержит потенциалзависимые Са2+-каналы (глава 2 I В I б (2) (в)). При деполяризации мембраны каналы открываются и ионы Ca2+ входят в терминаль, запуская в активных зонах экзоцитоз нейромедиатора.
а. Роль Ca[‡‡]*. Слияние синаптических пузырьков с пресинаптической мембраной происходит при увеличении концентрации Ca2+ в цитозоле нервной терминали.
Синаптотагмин — белок синаптического пузырька, связывающийся с Ca2+ и регулирующий экзоцитоз. Синаптотагмин участвует также в реорганизации при- мембранного цитоскелета, что важно для секреции медиатора.
б. Узнавание (рис. 8-5). Предшествующий слиянию синаптических пузырьков и плазмолеммы процесс узнавания синаптическим пузырьком пресинаптической мембраны происходит при взаимодействии мембранных белков (синаптобревин, SNAP-25 и синтаксин).
в. Активные зоны. В пресинаптической мембране выявлены т.н. активные зоны — участки утолщения мембраны, в которых происходит экзоцитоз. Активные зоны расположены против скоплений рецепторов в постсинаптической мембране, что уменьшает задержку в передаче сигнала, связанную с диффузией нейромедиатора в синаптической щели.
г. Влияние токсинов. Синтаксин, SNAP-25 и синаптобревин — мишени ботулиничес- кого токсина, необратимо подавляющего слияние синаптических пузырьков с пресинаптической мембраной. Мишень столбнячного токсина — синаптобревин.
- Постсинаптическая часть. Постсинаптическая мембрана содержит рецепторы нейромедиатора, ионные каналы.
- Синаптическая передача — сложный каскад событий. Многие неврологические и психические заболевания сопровождаются нарушением синаптической передачи. Различные лекарственные препараты влияют на синаптическую передачу, вызывая нежелательный эффект (например, галлюциногены) или, наоборот, корригируя патологический процесс (например, психофармакологические средства [антипсихотические препараты]),
а. Механизм. Синаптическая передача возможна при реализации ряда последовательных процессов: синтеза нейромедиатора, его накопления и хранения в синаптических пузырьках вблизи пресинаптической мембраны, высвобождения нейромедиатора из нервной терминали, кратковременного взаимодействия нейромедиатора с рецептором, встроенным в постсинаптическую мембрану, разрушения нейромедиатора или захвата его нервной терминалью.
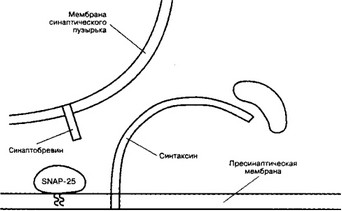
Рис. 8-5. Белки, участвующие в слиянии синаптических пузырьков с пресинаптической мембраной. Синаптобревин, встроенный в мембрану синаптического пузырька, взаимодействует с белками пресинаптической мембраны (SNAP-25 и синтаксин) [из Hata Yet al, 1993]
(б) Тормозные синапсы. При гиперполяризации возбудимость мембраны уменьшается, и потенциалы действия не генерируются.
- Удаление нейромедиатора из синаптической щели происходит двояко: инактивацией ферментом, захватом терминалью.
(а) Инактивация нейромедиатора. Кратковременность взаимодействия нейромедиатора с рецептором достигается разрушением нейромедиатора специальными ферментами (например, ацетилхолина — ацетилхолинэстеразой).
(б) Захват нейромедиатора. В большинстве синапсов передача сигналов прекращается вследствие быстрого захвата нейромедиатора пресинаптической терминалью.
Транспортёры. Захват норадреналина осуществляют специфические Na+- и СГ-тран-
спортирующие белки (например, норадреналин-транспортирующий белок I) — мишени трициклических антидепрессантов (например, дезипрамин и имипра- мин). Система захвата биогенных аминов — точка приложения антидепрессантов и таких препаратов, как кокаин и амфетамины. Дефекты транспортёров норадреналина и серотонина — кандидаты на роль первопричины при психических расстройствах, таких, как маниакально-депрессивные состояния.
б. Нейромедиаторы
- Химия. Большинство нейромедиаторов — аминокислоты и их производные. Одни нейроны модифицируют аминокислоты с образованием аминов (норадреналин, серотонин, ацетилхолин), другие — нейромедиаторов пептидной природы (эндор- фины, энкефалины). Лишь небольшое количество нейромедиаторов образовано не аминокислотами. Нейроны могут синтезировать более одного нейромедиатора.
- Наиболее распространённые нейромедиаторы (табл. 8-1)
(а) Ацетилхолин секретируется из терминалей соматических мотонейронов (нервно-мышечные синапсы), преганглионарных волокон, постганглионарных хо- линергических (парасимпатических) волокон вегетативной нервной системы и разветвлений аксонов многих нейронов ЦНС (базальные ганглии, двигательная кора). Синтезируется из холина и ацетил-КоА при помощи холинацетилтран- сферазы, взаимодействуете холинорецепторами нескольких типов. Кратковременное взаимодействие лиганда с рецептором прекращает ацетилхолинэстера- за, гидролизующая ацетилхолин на холин и ацетат.
- Болезнь Альцхаймера. При этом заболевании происходит гибель нейронов (в т.ч. холинергических) в коре мозга и гиппокампе.
(H) Отравления
- Ботулизм. Токсин Clostridium botulinum угнетает секрецию ацетилхолина.
Таблица 8-1. Нейромедиаторы
Нейропептиды
Аминокислоты
у-аминомасляная кислота
глицин
глутамат
N-метил-О-аспартат (NMDA)
Моноамины
адреналин
дофамин
норадреналин
серотонин
- Фосфорорганические соединения ингибируют ацетилхолинэстеразу, что приводит к увеличению количества ацетилхолина в синаптической щели. При отравлении пралидоксим способствует отделению соединения от фермента, атропин защищает холинорецепторы от взаимодействия с избыточным количеством нейромедиатора.
- Бледная поганка. Токсины Amanita phalloides не только ингибируют активность ацетилхолинэстеразы, но и блокируют холинорецепторы,
(б) Дофамин — нейромедиатор в окончаниях некоторых аксонов периферических нервов и многих нейронов ЦНС (чёрное вещество, средний мозг, гипоталамус). После секреции и взаимодействия с рецепторами дофамин активно захватывается пресинаптической терминалью, где его расщепляет моноаминок- сидаза. Дофамин метаболизирует с образованием ряда веществ, в т.ч. гомованилиновой кислоты.
- Шизофрения. При этом заболевании наблюдается повышенная реактивность дофаминергической системы, что связывают с увеличением количества D2-pe- цепторов дофамина. Антипсихотические средства снижают активность дофаминергической системы до нормального уровня.
- Хорея наследственная — нарушение функции нейронов коры и полосатого тела, сопровождается повышенной реактивностью дофаминергической системы.
(iii) Болезнь Паркинсона — патологическое уменьшение количества нейронов в чёрном веществе и других областях мозга с уменьшением уровня дофамина и метионин-энкефалина, преобладанием эффектов холинергической системы. Применение i-ДОФА увеличивает уровень дофамина, амантадин стимулирует секрецию дофамина, бромокриптин активирует рецепторы дофамина. Антихоли- нергические препараты уменьшают активность холинергической системы мозга.
(в) Норадреналин секретируется из большинства постганглионарных симпатических волокон и является нейромедиатором между многими нейронами ЦНС (например, гипоталамус, голубоватое место). Образуется из дофамина путём гидролиза при помощи дофамин-р-гидроксилазы. Норадреналин хранится в синаптических пузырьках, после высвобождения взаимодействует с адре- норецепторами, реакция прекращается в результате захвата норадреналина пресинаптической частью. Уровень норадреналина определяется активностью тирозин гидроксилазы и моноаминоксидазы. Моноаминоксидаза и катехол-O- метилтрансфераза переводят норадреналин в неактивные метаболиты (норме- танефрин, З-метокси-4-гидрокси-фенилэтиленгликоль, З-метокси-4-гидроксимин- дальная кислота).
Норадреналин — мощный вазоконстриктор, эффект происходит при взаимодействии нейромедиатора с ГМК стенки кровеносных сосудов.
(г) Серотонин (5-гидрокситриптамин) — нейромедиатор многих центральных нейронов (например, ядра шва). Предшественником служит триптофан, гид- роксилируемый триптофангидроксилазой до 5-гидрокситриптофана с последующим декарбоксилированием декарбоксилазой 1-аминокислот. Расщепляется моноаминоксидазой с образованием 5-гидроксииндолуксусной кислоты.
- Депрессия характеризуется снижением количества двух нейромедиаторов (норадреналина и серотонина) и увеличением экспрессии их рецепторов. Антидепрессанты уменьшают число этих рецепторов.
- Маниакальный синдром. При этом состоянии увеличивается уровень норадреналина на фоне снижения количества серотонина и адренорецепторов. Литий снижает секрецию норадреналина, образование вторых посредников и увеличивает экспрессию адренорецепторов.
- Аутизм. Гиперсеротонинемия, но в 30-50% случаев без явных нарушений обмена серотонина в мозге.
(д) у-Аминомасляная кислота — тормозной нейромедиатор в ЦНС (базальные ганглии, мозжечок). Образуется из глутаминовой кислоты под действием декарбоксилазы глутаминовой кислоты, захватывается из межклеточного пространства пресинаптической частью и деградирует под влиянием трансаминазы у-амино- масляной кислоты.
0) Эпилепсия — внезапные синхронные вспышки активности групп нейронов в разных областях мозга, связывают со снижением тормозного действия у-ами- номасляной кислоты. Фенитоин стабилизирует плазмолемму нейронов и снижает избыточную секрецию нейромедиатора, фенобарбитал повышает связывание у-аминомасляной кислоты с рецепторами, вальпроевая кислота увеличивает содержание нейромедиатора.
- Состояние тревоги — психическая реакция, связанная с уменьшением тормозного эффекта у-аминомасляной кислоты. Бензодиазепины стимулируют взаимодействие нейромедиатора с рецептором и поддерживают ингибиторное действие у-аминомасляной кислоты.
(е) р-Эндорфин — нейромедиатор полипептидной природы многих нейронов ЦНС (гипоталамус, миндалина мозжечка, таламус, голубоватое место). Проопиоме- ланокортин транспортируется по аксонам и расщепляется пептидазами на фрагменты, одним из которых является p-эндорфин. Нейромедиатор секрети- руется в синапсе, взаимодействует с рецепторами на постсинаптической мембране, а затем гидролизуется пептидазами.
(ж) Метионин-энкефалин и лейцин-энкефалин — небольшие пептиды (5 аминокислотных остатков), присутствующие во многих нейронах ЦНС (бледный шар, таламус, хвостатое ядро, центральное серое вещество). Как и эн- дорфин, образуются из проопиомеланокортина. После секреции взаимодействуют с пептидергическими (опиоидными) рецепторами.
(з) Динорфины. Эта группа нейромедиаторов состоит из 7 пептидов близкой аминокислотной последовательности, которые присутствуют в нейронах тех же анатомических областей, что и энкефалинергические нейроны. Образуются из продинорфина, инактивируются путём гидролиза.
(и) Вещество P — нейромедиатор пептидной природы в нейронах центральной и периферической нервной системы (базальные ганглии, гипоталамус, спинномозговые узлы).
Боль. Передача болевых стимулов реализуется при помощи вещества P и опиоидных пептидов (глава 8.3 I I в (2) (6)).
(к) Глицин, глутаминовая и аспарагиновая кислоты. Эти аминокислоты в некоторых синапсах являются нейромедиаторами (глицин во вставочных нейронах спинного мозга, глутаминовая кислота — в нейронах мозжечка и спинного мозга, аспарагиновая кислота — в нейронах коры). Глутаминовая и аспарагиновая кислоты вызывают возбуждающие ответы, а глицин — тормозные.
(л) Другие нейромедиаторы (VIP, адреналин, бомбезин, брадикинин, вазоп- рессин, карнозин, нейротензин, соматостатин, холецистокинин). Их роль для синаптической передачи остаётся неясной. В синаптической передаче, возможно, участвует прион.
Д. Цитоскелет нейронов состоит из микротрубочек, промежуточных филаментов (нейрофи- ламенты) и микрофиламентов.
- Микротрубочки — наиболее крупные элементы цитоскелета, их диаметр 24 нм. С ними связывают внутриклеточный, в т.ч. аксонный, транспорт (глава 8.2 I Б 2). От перикариона по отросткам перемещаются различные вещества (белки, нейромедиаторы и т.д.), органеллы (митохондрии, элементы цитоскелета, везикулы и т.д.). Микротрубочки в перикарионе и дендритах (в отличие от аксона) не имеют направленной ориентации.
а. Ориентация (рис. 8-6). Большинство микротрубочек аксона (+)-концом направлено к терминали, а (-)-концом — к перикариону. Характер ориентации микротрубочек имеет важное значение для распределения по отросткам различных органелл. К (+)- концу перемещаются митохондрии и секреторные пузырьки, а к (-)-концу — рибосомы, мультивезикулярные тельца, элементы комплекса Гольджи.
б. т-Белок — один из белков, связанных с микротрубочками большинства клеточных типов. т-Белок связывается с тубулином и стимулирует сборку микротрубочек, образует между ними поперечные сшивки. Модифицированная форма т-белка, формирующего волокна из пары спиральных нитей в составе плотных аномальных структур (нейро- фибриллярных клубков), обнаружена в нейронах мозга при болезни Алъцхаймера.
- Нейрофибриллы. При импрегнации солями серебра в нейронах можно обнаружить сплетения нитевидных структур, в аксонах расположенных параллельно друг другу. Нейрофибриллы — типичный артефакт, образующийся при осаждении серебром белков цитоскелета.
- Микрофиламенты (см. главу 2 III Б 3).
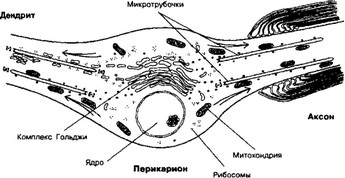
Рис. 8-6. Ориентация микротрубочек в отростках нейрона. Аксон отличается от дендритов полярной ориентацией микротрубочек. В нём микротрубочки своими (+)-концами направлены к терминали, а (-)-концами — к перикариону [из Black ММ, Baas PW, 1989)