
Морфогенетические процессы, реализация которых приводит к становлению жёстко организованной сложной системы, где каждый элемент индивидуален и знает своё место, — эта невообразимо (для осознания человеком) сложная задача всё же выполняется в нейроонтогенезе каждого человека. Ошибки программы (дефектные гены) или её реализации при выполнении этой задачи либо элиминируют развивающийся эмбрион (по разным оценкам, летали этого типа достигают 25%), либо приводят к появлению дефектов развития, в т.ч. нервно-психической сферы, и развитию пограничных состояний (например, IQ lt; 90). Жёсткость организации мозга определяют два момента; адресная миграция клеток и направленный рост их отростков. Феномены индукции, адресной миграции клеток, направленного роста аксонов, гибели клеток, т.н. нейротрофические взаимодействия, хорошо известны для нервной системы.
А. Индукция (см. главу 3 I В 2)
- Первичная эмбриональная индукция, приводящая к появлению нейроэктодермы, рассмотрена в главе 3 VI Б.
- Индукция хрусталика под влиянием глазного пузыря рассмотрена в главе 8.4.
- Индукционные межклеточные взаимодействия в нейроонтогенезе определяют судьбу каждого конкретного клеточного типа в нервной системе.
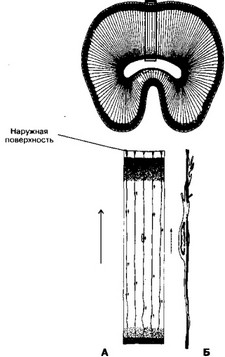
 Б. Адресная миграция клеток (рис. 8-2). В ходе морфогенеза мозга нейробласты выселяются из эпёндимного в мантийный слой (см. 8.1 I А). При формировании коры больших полушарий головного мозга и мозжечка нейробласты выселяются из эпёндимного слоя не только в мантийный слой, но также и в краевую вуаль (рис. 8-1). В мозжечке они формируют слой клеток Пуркинье. He все нейробласты этой локализации дифференцируются здесь в ганглиозные нейроны мозжечка. Часть из них мигрирует в обратном направлении, образуя зернистые клетки и клетки Гольджи II типа.
Б. Адресная миграция клеток (рис. 8-2). В ходе морфогенеза мозга нейробласты выселяются из эпёндимного в мантийный слой (см. 8.1 I А). При формировании коры больших полушарий головного мозга и мозжечка нейробласты выселяются из эпёндимного слоя не только в мантийный слой, но также и в краевую вуаль (рис. 8-1). В мозжечке они формируют слой клеток Пуркинье. He все нейробласты этой локализации дифференцируются здесь в ганглиозные нейроны мозжечка. Часть из них мигрирует в обратном направлении, образуя зернистые клетки и клетки Гольджи II типа.- Радиальная глия. Важное значение в направленной миграции клеток в пределах нервной трубки имеют специальные поддерживающие клетки радиальной глии, возникающие в раннем нейроонтогенезе. Их тела расположены в эпёндимном слое, а длинные отростки проходят через все слои нервной трубки до её наружной поверхности. По отросткам радиальной глии перемещаются нейробласты из эпёндимного слоя в наружные слои нервной трубки.
- Агирия — дефект развития в виде слабой выраженности извилин коры мозга вследствие искажённой миграции нейробластов в эмбриогенезе.
В. Направленный рост аксонов, как и адресная миграция клеток, осуществляется в рамках концепции «сигнал-ответ». Эта концепция объясняет, как нейрон узнаёт свою область иннервации и находит своего, часто единственного среди множества, клеточного партнёра, и как в развивающемся мозге многочисленные переплетающиеся отростки нейронов устанавливают связи с замечательной точностью. Отросток нейрона — аксон — сразу и без ошибок находит свои мишени. Направленный рост аксонов осуществляет конус роста.
- Конус роста — специализированная структура терминали растущего аксона, впервые детально описанная С.Рамон-и-Кахалем, имеет на конце булавовидное утолщение (ламел- лоподия), от которого отходят тонкие пальцевидные отростки — филоподии.
Филоподии растут в различных направлениях и исследуют потенциальное пространство роста аксона.
- Механизмы. На этот счёт существует несколько предположений. С.Рамон-и-Кахаль сформулировал представление о хемотропизме. Современный вариант этого представления — заранее размеченные пути.
а. Хемотропизм. Согласно этому представлению, рост аксонов происходит по градиенту концентрации специфических химических факторов, вырабатываемых в мишенях. Действительно, in vitro градиенты фактора роста нервов (NGF) и других веществ (например, ацетилхолина) влияют на направление роста аксонов.
б. Меченые пути (по другой терминологии, верстовые столбы) образованы молекулярными метками (своего рода знаками навигационной обстановки), закономерно распределёнными в потенциальном пространстве роста аксонов. По мере роста пионерский аксон последовательно считывает одну за другой метки, расположенные в межклеточном пространстве или на поверхности клеток, и растёт в нужном направлении. Вслед за ним мигрируют отростки других аксонов, совокупность которых формирует тракты в ЦНС и нервы на периферии. Ключевой момент представления о заранее размеченных путях — узнавание — обеспечивают молекулы адгезии.
в. Молекулы адгезии встроены в плазмолемму и расположены во внеклеточном матриксе.
- Мембранные молекулы ламеллоподии и филоподий взаимодействуют с комплементарными молекулами в пространстве роста и обеспечивают фиксацию конуса роста на поверхности мишени в нужном месте и в нужное время. Нейромодулин — одна из таких молекул.
- Молекулы внеклеточного матрикса: ламинин, фибронектин, коллаген. Наибольшее значение имеет ламинин. Этот крупномолекулярный белок базальной мембраны служит своего рода посредником между поверхностью клетки и молекулами внеклеточного матрикса (например, коллагеном IV типа). In vitro ламинин поддерживает адгезию, распластывание и миграцию клеток различных типов, рост аксонов, выживание нейронов и прикрепление конусов роста к субстрату.
Г. Физиологическая гибель клеток. Запрограммированная массовая смерть нейронов происходит на строго определённых этапах онтогенеза. Естественная гибель нейронов прослежена как в ЦНС, так и в периферической нервной системе. Объём субпопуляции гибнущих нейронов оценивают в широком интервале — от 25 до 75%. Иногда в популяции погибают все нейроны (например, несущие метку для направленного роста аксонов).
Причины гибели нейронов объясняют по-разному.
а. Преимущество в борьбе за выживание имеют те нейроны, которые раньше появляются, быстрее растут, образуют более эффективные контакты с мишенями, менее зависимы от трофических факторов, способны быстро увеличивать число рецепторов для этих факторов, наконец, имеют более эффективный ретроградный аксонный транспорт (см. главу 8.2 I Б 2) трофических факторов. Другими словами, преимущество в выживании имеют нейроны, наиболее активно участвующие в межклеточных взаимодействиях.
б. Выживание некоторых нейронов беспозвоночных полностью контролируется. Так, у почвенной нематоды мутации по гену ced-З предотвращают запрограммированную гибель нейронов. Найдены также гены, ответственные за выбор между жизнью и смертью клеток. Аналоги таких генов имеются и у человека.
в. Причины постоянной гибели нейронов (её именуют физиологической) в постнаталь- ном онтогенезе не очень понятны. В среднем у человека за год погибает около 10 млн. клеток, а в течение жизни мозг теряет около 0,1% всех нейронов. Бытует наиболее поверхностное объяснение — влияние неблагоприятных факторов микроокружения (например, сбои в кровоснабжении) и внешней среды.
Д. Нейротрофические взаимодействия. Эта концепция предусматривает информационный обмен между нейронами, отличающийся от химической передачи возбуждения в синапсах. Согласно концепции, такой обмен поддерживает фенотип взаимодействующих клеток на уровне, адекватном выполнению их функции (см., например, нейротрофический эффект нерва на мышцу в главе 7 I E I).
Е. Регенерация. Нейроны относят к статической клеточной популяции. Это означает, что в постнатальном онтогенезе не происходит образования новых нейронов. Следовательно, погибающие нейроны не восстанавливаются. Ho из этого не следует, что в нервной системе отсутствует регенерация. Она осуществляется за счёт восстановления целостности повреждённых нейронов, роста их отростков, размножения глиальных и шванновских клеток. Всё это, а также изменение связей между регенерирующими и интактными клетками при благоприятных условиях могут привести к восстановлению функции в нервной системе.
Ж. Трансплантация. При болезни Паркинсона дегенерация нейронов чёрной субстанции (substantia nigra) вызывает дефицит дофамина в области стриатума и нарушения в центрах и путях двигательного контроля. Интрацеребральная трансплантация содержащих дофамин клеток — приемлемая альтернатива при недостаточно эффективной фармакотерапии.