
В самой сжатой форме все появившиеся со времен П. Эрлиха гипотетические построения, касающиеся феномена иммунологической специфичности, можно разбить на две группы: инструктивные и селективные.
Инструктивные теории рассматривали антиген в качестве пассивного материала — матрицы, на которой формируется анти- генсвязующий участок антител. По этой теории все антитела имеют одну и ту же последовательность аминокислотных остатков. Различия касаются третичной структуры и возникают в процессе окончательного формирования молекулы антитела вокруг антигена. В настоящее время инструктивные теории полностью оставлены и имеют лишь исторический интерес. Они не выдержали проверки временем и вошли в противоречие с данными как иммунологии, так и молекулярной биологии. С иммунологических позиций они не объясняли, во-первых, почему количество антител в молярном отношении значительно больше количества проникшего в организм антигена, и, во-вторых, не отвечали на вопрос, за счет чего формируется иммунологическая память. С позиций молекулярной биологии они противоречили основной догме биоло-
о
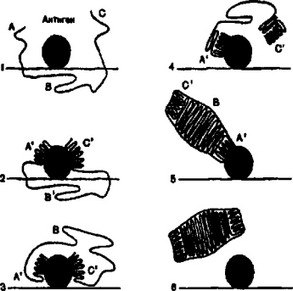
Рис. В.7. Инструктивная теория.
По этой теории все антитела имеют одну и ту же последовательность аминокислотных остатков. Различия касаются третичной структуры и возникают в процессе окончательного формирования молекулы антитела вокруг антигена; А, В, С — участки полипептидной цепи. Теория противоречит современным фактам иммунологии и молекулярной биологии и представляет только исторический интерес
гии, гласящей, что специфичность белка строго закодирована в последовательности нуклеотидов ДНК в хромосоме, а третичная структура связана с первичной структурой и не может в определенных пределах свободно меняться под влиянием факторов внешней среды без нарушения или полной потери функции.
Более плодотворными оказались селективные теории вариабельности антител. История развития иммунологаческой мысли в направлении селективных теорий началась с первого построения П.Эрлиха (см. выше). В основе всех селективных теорий лежит
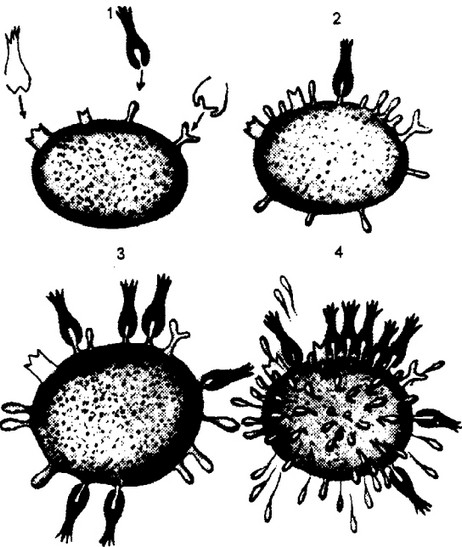
Рис. В.8. Теория “боковых целей” П.Эрлиха.
Данная теория является первой селективной теорией иммунитета. На поверхности клетки, способной образовывать антитела, имеются комплементарные к введенному антигену структуры — “боковые цепи”. Взаимодействие антигена с “боковой цепью” приводит к ее блокаде и как следствие — к компенсаторному повышенному синтезу и выходу в межклеточное пространство соответствующих “цепей”, выполняющих функцию антител
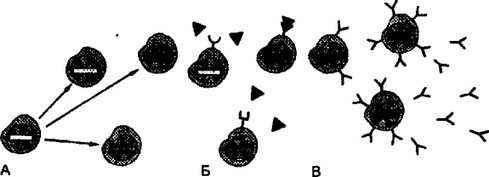
Рис. В.9. Клонально-селекционная теория М.Ф.Бернета.
При дифференциронке лимфоцитов от стволовой кроветворной клетки и при параллельно идущем процессе мутационных изменений б генах, ответственных за синтез антител, возникают клоны клеток (А), которые способны взаимодействовать с антигеном одной конкретной специфичности (Б). В результате подобного взаимодействия формируется отобранный по специфичности клон, который либо секретирует антитела заданной специфичности, либо обеспечивает клеточную реакцию (В)
представление о том, что специфичность антител предопределена, и антиген выступает лишь в качестве фактора отбора соответствующих по специфичности иммуноглобулинов.
В 1955 г. вариант селективной теории выдвинул Н.Ерне (1955). По его представлениям, в организме постоянно присутствуют антитела самой разнообразной специфичности. Антитело после взаимодействия с соответствующим антигеном поглощается фагоцитирующими мононуклеарами, что приводит к активной продукции этими клетками антител исходной специфичности.
Особое место в иммунологии занимает клонально-селекционная теория иммунитета М. Бернета (1059). Он использовал представления П. Эрлиха и Н. Ерне о предсуществовании антител разной специфичности, но указывал на то, что каждое специфическое антитело синтезируется отдельным клоном клеток. По М. Бер- нету, при дифференцировке лимфоцитов от стволовой кроветворной клетки и при параллельном процессе мутационных изменений в генах, контролирующих синтез специфических антител, возникают клоны клеток, которые способны взаимодействовать только с антигеном соответствующей специфичности. В результате такого взаимодействия формируется отобраный по специфичности клон, который либо секретирует антитела заданной специфичности, либо обеспечивает строго специфическую клеточную реакцию. Клонально-селекционный принцип организации иммунной системы, выдвинутый М. Бернетом, полностью подтвердился в настоящее время. Недостатком теории являются представления о том, что многообразие антител возникает только за счет мутационного процесса.
В то время, когда М. Вернет разрабатывал свою теорию, ничего не было известно о генах иммуноглобулинов и их реком-би- нации в процессе созревания В-клеток. Основной принцип селекции специфических клонов сохранен в теории зародышевой линии Л. Худа и соавт. (1971). Однако первопричину многообразия клонов авторы видят не в повышенной мутабельности иммуноглобулиновых генов, а в исходном зародышевом их предсуществовании. Весь набор V-генов, контролирующих вариабельную область иммуноглобулинов, представлен изначально в геноме и передается от поколения к поколению без изменений. В процессе развития В-клеток происходит рекомбинация иммуноглобулиновых генов,
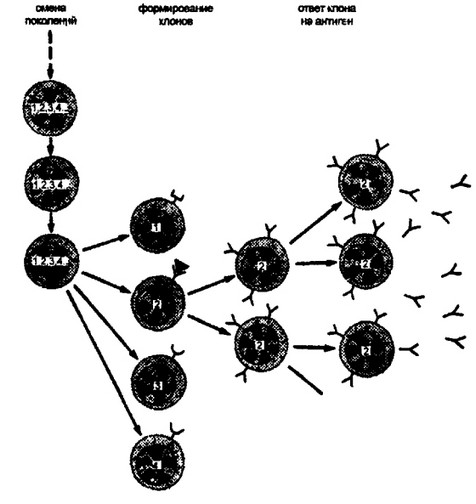
Рис. В.10. Теория зародышевой линии JLXyaa и др.
Весь набор V-генов (1. 2, 3, 4 ...), контролирующих вариабельную область иммуноглобулинов, представлен в геноме и передается от поколения к поколению без изменений. В процессе созревания В-клеток при индивидуальном развитии активируется один из V- генов, который в результате взаимодействия с С- геном кодирует иммуноглобулин определенной специфичности. Продукция строго специфических антител (иммуноглобулинов) есть функция определенного В-кле- точного клона
так что отдельно взятая созревающая В-клетка способна синтезировать иммуноглобулин одной специфичности. Такая моноспеци- фическая клетка становится источником клона В-клеток, продуцирующих определенный по специфичности иммуноглобулин.
Таким образом, теоретическая мысль в иммунологии развивалась от представлений П. Эрлиха о предсуществовании специфических антител и выдвижения клонально-селекционного принципа М. Бернетом до представлений о том, что клональность развития В-клеток предопределена в зародышевой линии и формируется в результате рекомбинации иммуноглобулиновых генов. Объединяющим моментом всех этих теоретических построений является убежденность в том, что антиген — лишь фактор селекции, но не участник формирования специфичности.