
Стадии развития от стволовой кроветворной клетки до незрелого В-лимфоцита проходят на территории костного мозга под прямым воздействием стромального микроокружения. Простым доказательством подобного утверждения являются опыты in vitro. Удаление из клеточной культуры стромальных элементов прерывает формирование В-лимфоцитов из стволовых клеток костного мозга. Реконструкция культуры восстанавливает процесс накопления В-клеток.
Участие клеток стромы в дифференцировке В-лимфоцитов проявляется в двух формах. Ранние этапы развития В-лимфоцитов зависят от прямого контактного взаимодействия со стромальными элементами. На более поздних этапах В-лимфоциты испытывают воздействие гуморальных факторов.
Взаимодействие наиболее ранних предшественников В-кле- ток со стромальными клеточными элементами осуществляется с помощью поверхностных адгезивных молекул (рис. 8.1). Первые контактные отношения возникают на уровне стволовой кроветворной клетки и общего лимфоидного предшественника через взаимодействие между гиалуроновой кислотой клеточной поверхности стромальных клеток и CD44. Это типичный адгезивный белок с относительно небольшой мол. массой 80-95 кД экспрессируется на поверхности различных типов лейкоцитов и эритроцитов. Помимо участия в установлении физического контакта между клетками стромы и ранними предшественниками В-клеток он задействован и в других формах межклеточных взаимоотношений, а также в процессах клеточной мифации и метастаэирования. Вероятно,связывание CD44 с лигандом не несет какой-либо прямой сигнальной функции, но стимулирует взаимодействие рецептора c-kit ранних про-В-клеток с другим рецептором стромальных клеток — SCF (сокращение от англ, “stem-cell factor”). Молекула c-kit обладает свойствами тирозинкиназ. В результате связывания этих поверхно-
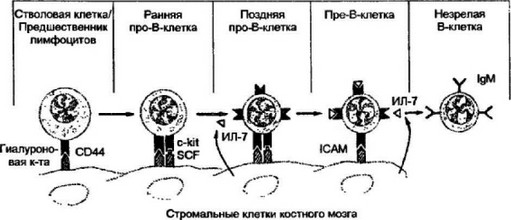
Рис. 8.1. Диференцяровка В-клеток н костном мозге.
Весь путь развития В-клеток в костном мозге делят на два основных этапа. Первый из них проходит при доминирующем участии адгезивных молекул: CD44, c-kit и SCF. На этом этапе стволовая кроветворная клетка диффсрс наиру стен в лимфоцитарный предшественник, общий для Т- и В-, от которого образуется ранняя про-В-клетка. На втором этапе в процесс костномозговой диффсрснии- ровки включаются цитокины, и в первую очередь интерлейкин-7 (ИЛ-7). В результате путь развития от ранней про-В-клетки идет через образование поздней про- В-клетки, которая в свою очередь дифференцируется в пре-В-клстки. Заключительной клеточной формой дифференциронки в костном мозге является незрелая В-клетка. Отличительными чертами незрелой Вв-клстки являются экспрессия на клеточной поверхности IgM, но отсутствие IgD, который появляется позднее у зрелых В-клеток периферии
стных молекул происходит активация c-kit, что влечет за собой усиление пролиферации и переход В-клсточных предшественников на более продвинутый уровень дифференцировки — стадию поздних про-В-клеток.
Этот этап дифференцировки характеризуется появлением на поверхности поздних В-клеток рецептора к интерлсйкину-7 и включением в процесс соответствующего цитокина, секретируемого стромальными клетками. Активность интерлейкина-7 в качестве ростового фактора вполне достаточна для поддержания пролиферации и выживания развивающихся клеток. В результате их зависимость от SCF снижается и на стадии пре-В-клеток полностью пре кращается. Контакт пре-В-клеток со стромой осуществляют адге- зины 1САМ.
Процесс В-клеточного развития в костном мозге завершается формированием незрелой В-клстки, экспрессирующей поверхностный IgM и подготовленной к миграции на периферию.
Кроме отмеченных, имеется еще целый ряд рецепторов, позволяющих следить за развитием В-клеток (табл. 8.1). Функция этих поверхностных структур нс является прерогативой только развивающихся В-клсток. Они включены в процессы клеточного взаимодействия, представления антигена в иммуногенной форме и др. Некоторые их свойства отмечены н подписи к таблице 8.1.
Таблица 8.1
Изменение экспрессии молекул клеточной поверхности в процессе
дифференцировки В-клеток
CD45R — одна из изоформ общего лейкоцитарного антигена; экспрессируется на клетках различных Т- и В-субпопуляций, моноцитах, макрофагах; мол. масса 180 кД; трансмембранный белок, цитоплазматический домен которого обладает тирозинспе- цифической фосфатазной активностью и активирует тирозинки- назы, ассоциированные с антигенными рецепторами и корецеп- торами Т- и В-клеток, таким образом усиливая прохождение сигнала внутрь клетки от антигенрецепторного взаимодействия.
CD19 — экспрессируется на В-клетках; мол. масса — 95 кД; выполняет функцию корецептора у В-клеток.
CD40 — экспрессируется на В-клетках различной степени зрелости, моноцитах, дендритных клетках; мол. масса — 50 кД; выполняет функцию рецептора при костимуляции В-клеток.
CD20 — экспрессируется на В-клетках; мол. масса — 33-37 кД; возможно, участвует в активации В-клеток.
CD21 — экспрессируется на В-клетках, фолликулярных дендритных клетках; мол. масса — 145 кД; выполняет функцию рецептора для C3d - компонента комплемента и вируса Эпштейн-Барр; вместе с CD 19 и CD81 образует корецептор у В-клеток.
Участие клеток стромы в дифференцировке В-лимфоцитов проявляется в двух формах. Ранние этапы развития В-лимфоцитов зависят от прямого контактного взаимодействия со стромальными элементами. На более поздних этапах В-лимфоциты испытывают воздействие гуморальных факторов.
Взаимодействие наиболее ранних предшественников В-кле- ток со стромальными клеточными элементами осуществляется с помощью поверхностных адгезивных молекул (рис. 8.1). Первые контактные отношения возникают на уровне стволовой кроветворной клетки и общего лимфоидного предшественника через взаимодействие между гиалуроновой кислотой клеточной поверхности стромальных клеток и CD44. Это типичный адгезивный белок с относительно небольшой мол. массой 80-95 кД экспрессируется на поверхности различных типов лейкоцитов и эритроцитов. Помимо участия в установлении физического контакта между клетками стромы и ранними предшественниками В-клеток он задействован и в других формах межклеточных взаимоотношений, а также в процессах клеточной мифации и метастаэирования. Вероятно,связывание CD44 с лигандом не несет какой-либо прямой сигнальной функции, но стимулирует взаимодействие рецептора c-kit ранних про-В-клеток с другим рецептором стромальных клеток — SCF (сокращение от англ, “stem-cell factor”). Молекула c-kit обладает свойствами тирозинкиназ. В результате связывания этих поверхно-
Рис. 8.1. Диференцяровка В-клеток н костном мозге.
Весь путь развития В-клеток в костном мозге делят на два основных этапа. Первый из них проходит при доминирующем участии адгезивных молекул: CD44, c-kit и SCF. На этом этапе стволовая кроветворная клетка диффсрс наиру стен в лимфоцитарный предшественник, общий для Т- и В-, от которого образуется ранняя про-В-клетка. На втором этапе в процесс костномозговой диффсрснии- ровки включаются цитокины, и в первую очередь интерлейкин-7 (ИЛ-7). В результате путь развития от ранней про-В-клетки идет через образование поздней про- В-клетки, которая в свою очередь дифференцируется в пре-В-клстки. Заключительной клеточной формой дифференциронки в костном мозге является незрелая В-клетка. Отличительными чертами незрелой Вв-клстки являются экспрессия на клеточной поверхности IgM, но отсутствие IgD, который появляется позднее у зрелых В-клеток периферии
стных молекул происходит активация c-kit, что влечет за собой усиление пролиферации и переход В-клсточных предшественников на более продвинутый уровень дифференцировки — стадию поздних про-В-клеток.
Этот этап дифференцировки характеризуется появлением на поверхности поздних В-клеток рецептора к интерлсйкину-7 и включением в процесс соответствующего цитокина, секретируемого стромальными клетками. Активность интерлейкина-7 в качестве ростового фактора вполне достаточна для поддержания пролиферации и выживания развивающихся клеток. В результате их зависимость от SCF снижается и на стадии пре-В-клеток полностью пре кращается. Контакт пре-В-клеток со стромой осуществляют адге- зины 1САМ.
Процесс В-клеточного развития в костном мозге завершается формированием незрелой В-клстки, экспрессирующей поверхностный IgM и подготовленной к миграции на периферию.
Кроме отмеченных, имеется еще целый ряд рецепторов, позволяющих следить за развитием В-клеток (табл. 8.1). Функция этих поверхностных структур нс является прерогативой только развивающихся В-клсток. Они включены в процессы клеточного взаимодействия, представления антигена в иммуногенной форме и др. Некоторые их свойства отмечены н подписи к таблице 8.1.
Таблица 8.1
Изменение экспрессии молекул клеточной поверхности в процессе
дифференцировки В-клеток
|
Общий лредшествен ник Т- я В - ЛИНИН |
Ранняя про- В-клетка |
Поздняя про-В- клеткя |
Пре-В- клетка |
Незрелая В-клетка |
Зрелая В-клетка |
|
CEM5R |
CD45R |
CD45R |
CIM5R |
CD45R |
CD45R |
|
CDI9 |
CD19 |
CD19 |
CD19 |
CD19 |
CD19 |
|
CD38 |
CD38 |
CD38 |
CD38 |
|
|
|
|
СЕМО |
CD40 |
CD40 |
CD40 |
СЕМО |
|
|
|
|
CD20 |
CD20 |
CD20 |
|
|
|
|
|
CD21 |
CD2I |
|
I кл. II ГКГ |
кл. 11 ГКГ |
кл. II ГКГ |
кл. 11 ГКГ |
кл. 11 ГКГ |
|
|
|
|
|
й |
IgM |
IgM lgD |
CD45R — одна из изоформ общего лейкоцитарного антигена; экспрессируется на клетках различных Т- и В-субпопуляций, моноцитах, макрофагах; мол. масса 180 кД; трансмембранный белок, цитоплазматический домен которого обладает тирозинспе- цифической фосфатазной активностью и активирует тирозинки- назы, ассоциированные с антигенными рецепторами и корецеп- торами Т- и В-клеток, таким образом усиливая прохождение сигнала внутрь клетки от антигенрецепторного взаимодействия.
CD19 — экспрессируется на В-клетках; мол. масса — 95 кД; выполняет функцию корецептора у В-клеток.
CD40 — экспрессируется на В-клетках различной степени зрелости, моноцитах, дендритных клетках; мол. масса — 50 кД; выполняет функцию рецептора при костимуляции В-клеток.
CD20 — экспрессируется на В-клетках; мол. масса — 33-37 кД; возможно, участвует в активации В-клеток.
CD21 — экспрессируется на В-клетках, фолликулярных дендритных клетках; мол. масса — 145 кД; выполняет функцию рецептора для C3d - компонента комплемента и вируса Эпштейн-Барр; вместе с CD 19 и CD81 образует корецептор у В-клеток.