
Общеизвестна роль, которую играют онкогены в развитии опухолей. Утрата нормальной регуляции экспрессии протоонкогенов делает их активность бесконтрольной и превращает их в онкогены. Это происходит при пространственном перемещении протоонкогенов в процессе хромосомных
4.6. Опухоли иммунной системы — лимфопролиферативные...
перестроек или вследствие встраивания вирусных генов в геном клетки. Яркие и типичные примеры активации онкогенов, приводящие к развитию злокачественного процесса, обнаружены при лимфопролиферативных заболеваниях.
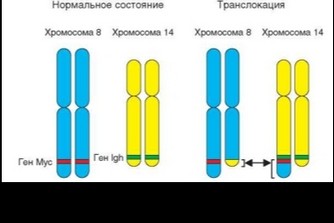
Подробно изучены транслокации участков хромосомы 8 (содержит протоонкоген c-myc) и хромосомы 18 (содержит протоонкоген bcl-2) в хромосому 14, в результате которых названные протоонкогены оказываются в непосредственной близости от гена Н-цепи иммуноглобулинов — IGH (рис. 4.41). Эти два типа перестроек служат предпосылкой развития соответственно В-клеточной лимфомы Беркита и других вариантов фолликулярной В-лимфомы. При лимфоме Беркита может происходить перенос фрагмента хромосомы 8 с геном также в хромосому 2 и хромосому 22, к локусам, занимаемым генами IGK и IGL, кодирующим соответственно к- и ^-цепи иммуноглобулинов. При той же лимфоме вслед за транслокацией участка, содержащего ген с-myc, может последовать транслокация участка, который содержит ген bcl-2, что повышает злокачественность лимфопролиферативного процесса.
Лимфома Беркита имеет вирусную природу. Ее вызывает вирус Эпштейна— Барр, часто присутствующий в организме человека в латентной форме или вызывающий инфекционный мононуклеоз. Это заболевание состоит во временной доброкачественной пролиферации В-лимфоцитов. Причина пролиферации В-клеток при этом заключается в том, что один из продуктов вируса является транскрипционным фактором, способствующим индукции генов, причастных к митогенезу. При этом не происходит транслокаций указанного выше типа. В тех случаях, когда происходит транслокация, развивается злокачественный лимфопролиферативный процесс — лимфома Беркита. Обычно такая транслокация происходит редко, лишь при условии действия кофакторов. Однако в некоторых регионах Африки, где эти кофакторы распространены, лимфома Беркита имеет характер эпидемии.
Тот же вирус при сочетании с действием других кофакторов вызывает два других заболевания — рак носоглотки (также имеющий эпидемический характер) и лимфогранулематоз. Таким образом, инфицирование вирусом Эпштейна—Барр — ведущее, но не единственное условие формирования генетических перестроек и развития лимфом. В то же время между формированием транслокаций и развитием лимфопролиферативного процесса есть тесная связь.
Уже упоминалось о том, что частота злокачественных процессов, связанных с пролиферацией В-клеток, у человека выше, чем частота Т-клеточных лимфопролиферативных процессов. Обратную ситуацию наблюдают при анализе злокачественного поражения лимфоидной ткани на фоне подавления иммунитета — при наследственных иммунодефицитах или после действия иммуносупрессивных агентов (облучения, химиопрепаратов). В этих случаях малигнизации чаще подвергаются Т-лимфоциты.
Особый интерес представляет атаксия-телеангиоэктазия (см. раздел 4.7.1.3), при которой иммунодефицит сочетается с нестабильностью хромосом. При этом заболевании выявляют 2 основных типа транслокаций, один из которых не сопровождается развитием опухолевых процессов, а другой приводит к формированию Т-клеточного лейкоза. Второй тип транслокаций затрагивает ген, локализованный в хромосоме 14 и обозначаемый TCL1 (по аналогии с геном BCL2). Чаще всего фрагмент, содержащий этот ген, перемещается в участки хромосом, в которых локализованы гены полипептидных цепей TCR, в первую очередь TCRA (14q11), кодирующий а/5-цепи TCR, несколько реже — к гену TCRB (7q35). В данном случае связь с вирусной инфекцией не установлена.
Вирус является основной причиной другого Т-лимфопролиферативно- го процесса — острого Т-клеточного лейкоза взрослых. Это заболевание вызывается вирусом HTLV-1 — лимфотропным вирусом, по ряду свойств сходным с ВИЧ. Как и ВИЧ, он обладает сродством к CD4+ Т-лимфоцитам, однако вызывает их трансформацию, а не разрушение. Как и при лимфоме Беркита, для индукции этого заболевания требуется «сотрудничество» вируса с кофакторами, имеющими эндемичную природу. Эндемичная зона для этой формы лейкоза — южные Японские острова.
Во всех рассмотренных случаях протоонкогены перемещаются в участки хромосомы, содержащие гены антигенраспознающих рецепторов BCR и TCR, в которых в норме происходят процессы реаранжировки. Очевидно, это не случайно и вирусы используют повышенную готовность этих участков к рекомбинации. Типичное следствие таких перемещений генов — злокачественная лимфопролиферация — находится в очевидной связи с запуском физиологической активации при воздействии на продукты этих генов — рецепторы для антигенов. Инфекционный мононуклеоз является процессом, моделирующим ту же ситуацию в доброкачественном варианте.
Наиболее четкий ответ на вопрос, способны ли малигнизированные лимфоциты выполнять присущие им в норме функции, получен при изучении малигнизированных плазмоцитов при множественной миеломе. Еще в начале 70-х годов было установлено, что при этом заболевании в сыворотке крови электрофоретически выявляют гомогенный пик (М-компо- нент), образуемый иммуноглобулинами. Затем было показано, что эти
иммуноглобулины обладают активностью антител со специфичностью, идентичной для всех молекул. Специфичность таких антител непредсказуема. Иногда эти антитела связывают даже синтетические гаптены. Однако чаще они направлены против иммунодоминантных компонентов бактерий, например, фосфорилхолина. Все эти данные свидетельствуют о моноклональном происхождении указанных иммуноглобулинов-антител. Это вполне соответствует представлениям о клональной структуре иммунной системы и служит свидетельством происхождения множественной миеломы в результате малигнизации клона плазмоцитов, «выбираемого» случайно.
Таким образом, по крайней мере при малигнизации, эффекторные клетки иммунной системы сохраняют свою функцию (в случае с плазмоцитом она состоит в секреции антител заданной специфичности). Однако на уровне популяции клеток и организма в целом, задача, связанная с иммунным ответом, при этом не выполняется, поскольку, во-первых, ответ малигнизированных плазмоцитов не является адаптивным, во-вторых, он моно- клонален, т.е. не может адекватно отразить множественность антигенных стимулов, проявляющихся при инфицировании и других видах биологической агрессии. Нормальная составляющая иммунного ответа при множественной миеломе страдает не только от ее «вытеснения» пролиферирующим клоном, но и от действия регуляторных механизмов. Они срабатывают в ответ на избыточное развитие моноклонального процесса и направлены на его ограничение, но в результате подавляют не его (этот процесс не реагирует на регуляторные сигналы), а нормальный иммунный ответ.
Моноклональность свойственна всем злокачественным лимфопролиферативным процессам. Однако в других случаях пролиферации зрелых форм лимфоцитов ее удается зарегистрировать не по продуктам, секрети- руемым клетками, а по результатам генетического анализа реаранжировки генов BCR и TCR. Характер перестройки генов идентичен для злокачественных клеток в каждом конкретном случае заболевания, но различен для каждого из них.
Представляют интерес немногочисленные результаты систематического анализа комплекса цитокинов, продуцируемых малигнизированными лимфоцитами, хотя четкой связи разновидностей Т-лимфом с определенным типом дифференцировки Т-клеток, вопреки первоначальным ожиданиям, установить не удается.
По-видимому, в большинстве злокачественных процессов с вовлечением зрелых лимфоцитов трансформированные клетки сохраняют сформировавшиеся функции, особенно секреторные, так же, как экспрессию свойственных им маркерных молекул, функционально значимых рецепторов и молекул адгезии. «Правильное» следование свойственным им путям миграции позволяет считать, что экспрессируемые молекулы действительно выполняют свои функции. В то же время сомнительно, чтобы малигнизированные клетки могли адекватно реагировать на регуляторные сигналы, контролирующие их специфическую активность, и практически невозможно проверить, способны ли лейкозные CD8+ Т-клетки осуществлять лизис специфических клеток-мишеней, поскольку специфичность их TCR никогда не удавалось установить.
Немногочисленные попытки оценить при Т-клеточных лимфопролиферативных заболеваниях состояние компонентов иммунной системы, не затронутых малигнизацией, свидетельствуют о преобладании отрицательных регуляторных воздействий. Последние обусловлены активацией регуляторных Т-клеток и направлены на ограничение неадекватной пролиферации злокачественных клонов Т-лимфоцитов. К этому надо добавить, что объем нормального компонента иммунной системы, как правило, уменьшен в результате его вытеснения малигнизированными Т-клетками. В результате при всех заболеваниях указанной группы (особенно при лейкозах) развивается тотальный иммунодефицит.