
NK-клетки развиваются в костном мозгу и происходят от того же общего лимфоидного предшественника CLP, который дает начало всем разновидностям лимфоцитов (рис. 2.32). В развитии естественных киллеров важную роль играет влияние микроокружения, реализуемое как через прямые межклеточные контакты, так и посредством цитокинов, например, взаимодействие представленного на мембране NK-клеток лимфотоксина а с рецепторами на стромальных клетках. Смесь цитокинов, содержащая IL-7 и IL-15, а также Flt-3-лиганд, необходима для дифференцировки естественных киллеров из костномозговых клеток-предшественников in vitro. На этапе выбора пути дифференцировки CLP в направлении Т- и В-линий, потенциал развития NK-клеток сохраняется за про-Т-клетками, иногда обозначаемыми как пре-Т/NK-клетки (Т/NKР). Развитие Т/NKР-клеток блокируют дефекты, затрагивающие гены мембранных молекул CD3e и
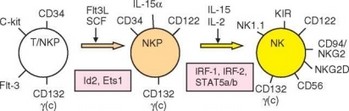
Рис. 2.32. Основные стадии развития естественных киллеров: общий предшественник NK- и T-лимфоцитов (T/NKP), специализированный предшественник NK-клеток (NKP) и зрелая NK-клетка. Указаны мембранные маркеры клеток; в прямоугольниках — дифференцировочные факторы, регулирующие соответствующую стадию развития
FceRIy (CD3e — компонент рецепторного комплекса Т-клеток; FceRIy в качестве передающей сигнал полипептидной цепи входит в состав ряда рецепторов, в том числе FceRI).
На следующем этапе клетки линии естественных киллеров окончательно отделяются от Т-линии. Это наиболее четко выражено при развитии тимоцитов — на самых ранних стадиях созревания (стадии тимоцитов DN1, DN2 — см. раздел 3.3.2.3) возможна их дифференцировка не только в Т-лимфоциты, но и в NK-клетки. Эта способность полностью утрачивается на стадии DN3-клеток, когда происходит перестройка V-генов TCR (начальные неспецифические этапы этой перестройки происходят и в предшественниках NK-клеток). Условие дифференцировки NK-клеток — экспрессия внутриклеточных факторов дифференцировки Id2 и Ets 1. Необходимые для дифференцировки Т-клеток факторы группы Notch блокируют развитие естественных киллеров.
После отделения от Т-линии дифференцирующиеся NK-клетки обозначают как пре-NK, или NKP-клетки (Natural killer progenitor). При переходе на эту стадию развития на поверхности клетки экспрессируется р-цепь (CD122), общая для рецепторов IL-2 и IL-15. К этому моменту другой компонент этих рецепторов — у(с)-цепь (от y-common), представляющая собой общую цепь для большой группы гемопоэтиновых рецепторов (см. раздел 2.5.5.2) уже представлена на клетках (она появляется на стадии NKTР). Вскоре клетка экспрессирует a-цепь рецептора для IL-15, и с этого момента IL-15 становится основным цитокином, определяющим дальнейшее развитие, выживаемость и гомеостаз NK-клеток.
Следующий этап развития, реализуемый при участии IL-15, IL-18 и IL-12, — формирование зрелой NK-клетки. Для этого этапа свойственно последовательное появление маркеров и рецепторов, характерных для NK-клеток: NK1.1, CD94/NKG2 и NKG2D. Важное условие экспрессии рецепторных молекул Ly49, CD94/NKG2 — взаимодействие NKР-клеток со стромальными клетками костного мозга, экспрессирующими молекулы МНС-I — лиганды этих рецепторов. В то же время на NK-клетках появляются интегрины [в частности р2-интегрин Мас-1 (аМР2)], и исчезает молекула
CD34. На последнем этапе созревания NK-клетка проходит несколько актов деления. IL-15-зависимая передача сигнала играет ключевую роль на этом этапе развития NK-клеток. Переход от стадии NKP к зрелым NK-клеткам блокируется при выключении генов самого IL-15, a-цепи его рецептора и связанной с ней тирозинкиназы Jak3, а также транскрипционных факторов STAT5 (a и b), обеспечивающих передачу сигнала от рецептора, и факторов IRF-1 и IRF-2, необходимых для секреции IL-15.
Зрелые NK-клетки покидают костный мозг и мигрируют в периферический отдел иммунной системы. Их расселение существенно отличается от распределения Т- и В-лимфоцитов. Значительная часть NK-клеток находится в циркуляции, составляя около 10% (5—15%) числа лимфоцитов периферической крови. Естественные киллеры содержатся в синусоидах печени, эндометрии матки и некоторых других солидных органах. Их содержание в лимфатических узлах и селезенке невелико (2,5—5%). В селезенке NK-клетки локализованы в основном в красной пульпе. Расселение естественных киллеров обусловено экспрессированными на их поверхности хемокиновыми рецепторами: CXCR4, CCR1 и CCR5. Эти рецепторы определяют миграцию клеток по градиенту хемокинов: CXCL12 (SDF-1) и многих Р-хемокинов. Только небольшая их часть (5% NK-клеток крови) экспрессирует рецептор CCR7, участвующий в преодолении эндотелиального барьера в посткапиллярных венулах и миграции лимфоцитов в Т-зоны вторичных лимфоидныех органов. На NK-клетках, кроме того, экспрессирован рецептор для фракталкина CX3CR.
Естественные киллеры — короткоживущие клетки (время полужизни составляет 7—10 сут). Как уже отмечалось, выживаемость NK-клеток зависит от IL-15. При отсутствии IL-15 они быстро погибают; при этом перенесенные NK-клетки не приживаются. IL-15 не действут на рецепторы NK-клеток напрямую. Этот цитокин предварительно связывают вспомогательные клетки и представляют его естественным киллерам, удлинняя действие и существенно повышая его эффект. Дополнительную роль в поддержании жизнеспособности NK-клеток играет IL-7. При снижении численности NK-клеток запускается механизм гомеостатической пролиферации, также обусловленный IL-15 и IL-7.
Рис. 2.32. Основные стадии развития естественных киллеров: общий предшественник NK- и T-лимфоцитов (T/NKP), специализированный предшественник NK-клеток (NKP) и зрелая NK-клетка. Указаны мембранные маркеры клеток; в прямоугольниках — дифференцировочные факторы, регулирующие соответствующую стадию развития
FceRIy (CD3e — компонент рецепторного комплекса Т-клеток; FceRIy в качестве передающей сигнал полипептидной цепи входит в состав ряда рецепторов, в том числе FceRI).
На следующем этапе клетки линии естественных киллеров окончательно отделяются от Т-линии. Это наиболее четко выражено при развитии тимоцитов — на самых ранних стадиях созревания (стадии тимоцитов DN1, DN2 — см. раздел 3.3.2.3) возможна их дифференцировка не только в Т-лимфоциты, но и в NK-клетки. Эта способность полностью утрачивается на стадии DN3-клеток, когда происходит перестройка V-генов TCR (начальные неспецифические этапы этой перестройки происходят и в предшественниках NK-клеток). Условие дифференцировки NK-клеток — экспрессия внутриклеточных факторов дифференцировки Id2 и Ets 1. Необходимые для дифференцировки Т-клеток факторы группы Notch блокируют развитие естественных киллеров.
После отделения от Т-линии дифференцирующиеся NK-клетки обозначают как пре-NK, или NKP-клетки (Natural killer progenitor). При переходе на эту стадию развития на поверхности клетки экспрессируется р-цепь (CD122), общая для рецепторов IL-2 и IL-15. К этому моменту другой компонент этих рецепторов — у(с)-цепь (от y-common), представляющая собой общую цепь для большой группы гемопоэтиновых рецепторов (см. раздел 2.5.5.2) уже представлена на клетках (она появляется на стадии NKTР). Вскоре клетка экспрессирует a-цепь рецептора для IL-15, и с этого момента IL-15 становится основным цитокином, определяющим дальнейшее развитие, выживаемость и гомеостаз NK-клеток.
Следующий этап развития, реализуемый при участии IL-15, IL-18 и IL-12, — формирование зрелой NK-клетки. Для этого этапа свойственно последовательное появление маркеров и рецепторов, характерных для NK-клеток: NK1.1, CD94/NKG2 и NKG2D. Важное условие экспрессии рецепторных молекул Ly49, CD94/NKG2 — взаимодействие NKР-клеток со стромальными клетками костного мозга, экспрессирующими молекулы МНС-I — лиганды этих рецепторов. В то же время на NK-клетках появляются интегрины [в частности р2-интегрин Мас-1 (аМР2)], и исчезает молекула
CD34. На последнем этапе созревания NK-клетка проходит несколько актов деления. IL-15-зависимая передача сигнала играет ключевую роль на этом этапе развития NK-клеток. Переход от стадии NKP к зрелым NK-клеткам блокируется при выключении генов самого IL-15, a-цепи его рецептора и связанной с ней тирозинкиназы Jak3, а также транскрипционных факторов STAT5 (a и b), обеспечивающих передачу сигнала от рецептора, и факторов IRF-1 и IRF-2, необходимых для секреции IL-15.
Зрелые NK-клетки покидают костный мозг и мигрируют в периферический отдел иммунной системы. Их расселение существенно отличается от распределения Т- и В-лимфоцитов. Значительная часть NK-клеток находится в циркуляции, составляя около 10% (5—15%) числа лимфоцитов периферической крови. Естественные киллеры содержатся в синусоидах печени, эндометрии матки и некоторых других солидных органах. Их содержание в лимфатических узлах и селезенке невелико (2,5—5%). В селезенке NK-клетки локализованы в основном в красной пульпе. Расселение естественных киллеров обусловено экспрессированными на их поверхности хемокиновыми рецепторами: CXCR4, CCR1 и CCR5. Эти рецепторы определяют миграцию клеток по градиенту хемокинов: CXCL12 (SDF-1) и многих Р-хемокинов. Только небольшая их часть (5% NK-клеток крови) экспрессирует рецептор CCR7, участвующий в преодолении эндотелиального барьера в посткапиллярных венулах и миграции лимфоцитов в Т-зоны вторичных лимфоидныех органов. На NK-клетках, кроме того, экспрессирован рецептор для фракталкина CX3CR.
Естественные киллеры — короткоживущие клетки (время полужизни составляет 7—10 сут). Как уже отмечалось, выживаемость NK-клеток зависит от IL-15. При отсутствии IL-15 они быстро погибают; при этом перенесенные NK-клетки не приживаются. IL-15 не действут на рецепторы NK-клеток напрямую. Этот цитокин предварительно связывают вспомогательные клетки и представляют его естественным киллерам, удлинняя действие и существенно повышая его эффект. Дополнительную роль в поддержании жизнеспособности NK-клеток играет IL-7. При снижении численности NK-клеток запускается механизм гомеостатической пролиферации, также обусловленный IL-15 и IL-7.