
наглядно демонстрирует роль органоразобщающих механизмов 2 и ПОН в развитии нарушений биологической целостности организма (БЦО) (рис 2.1).
Таблица 2.1
Интегративная медицина как самостоятельный раздел медицины
| Критерии науки | Содержание |
| Предмет
исследования |
Биологическая целостность организма (БЦО) |
| Объект
исследования |
Здоровый и больной человек |
| Цель | Сохранение БЦО человека в процессе его жизнедеятельности |
| Уровень
воздействия |
Индивидуальный многоуровневый: молекулярный, тканевой, органный, системный |
| Основное
направление практической деятельности |
Сохранение БЦО, руководствуясь принципом достаточности и взаимосоответсгвия энергетического статуса (ЭС) и функциональной состоятельности организма (ФСО) |
| Методолошческ ие основы | БЦО — это взаимосоответствующее единение ЭС и ФСО, способное удовлетворить основную биологическую потребность человека — динамическое самообновление |
| Основные
методы |
Биометрия для определения категорий БЦО, статусметрия для определения угрозоопасности нарушений ФСО, критичности ЭС, слабого звена ФСО, ведущей органной дисфункции в ЭС. Технологии профилактики преждевременной смерти. Технологии восстановления биоустойчивосги |
Представление об организме, как самовосстанавливающейся системе, требует нового методологического подхода к сепсисо- логии, которая, кроме этиологического, патофизиологического и клинического разделов, должна включать биологический.
Современная наука придает все большее значение изучению процессов биоэнергетики, ибо жизнедеятельность любого организма определяется его энергетическим статусом. Эту потребность в полной мере удовлетворяет интегративная медицина (ИМ), целью которой является сохранение биологической целостности организма в процессе его жизнедеятельности (табл. 2.1).
2 Любое действие практикующего врача должно быть направленным на сохранение биологической устойчивости организма пациента, руководствуясь принципом создания достаточности и взаимосоответствия энергетического статуса (ЭС) и функционального состояния организма (ФСО). БЦО характеризуется энергетическим статусом пациента, формирующимся в процессе его самовосстановления.
- Эти методологические предпосылки к пониманию биоустойчивости определяют коренную трансформацию и смену научных представлений и методов профессиональной деятельности. При обучении ИМ происходит переосмысление основ теории, изменение методологических предпосылок, стиля профессионального мышления и самой научной картины мира. Интегративная медицина использует технологии сохранения БЦО, руководствуясь принципом создания достаточности и взаимосоответствия энергетического статуса и функционального состояния организма.
Биоустойчивость определяется структурно-функциональным состоянием пациента, формирующимся в процессе его самосохранения. При этом характер ЭС оценивается по уровню биологического потенциала (БП) организма, отражающему многообразие проявлений биологической способности ЭС удовлетворить основные биологические потребности пациента, соответствующие интенсивности обмена веществ, потреблению энергосубстратов и кислорода. Биометрическим мерилом и эталоном величины БП служит его базальная величина, соответствующая индивидуальной способности ЭС гарантировать собственную биорегуляторную готовность к немедленному выполнению любых проявлений функциональной состоятельности организма (ФСО), сохраняя возможность динамического
самообновления. Адаптивноспособному состоянию ЭС соотве- 2 тствует энергоструктурный баланс с нормоэнергобиотическим уровнем БП. Спектр возможностей ЭС выражают сдвиги энер- гострукгурного баланса, которым соответствуют гипер-, гипо- и патобиотические виды БП.
ФСО — обязательная составляющая БЦО, которая определяет саму способность организма удовлетворить свои основные биологические потребности: рост, развитие, самообновление, размножение и, наконец, смерть. Поэтому ФСО, как главная биометрическая составляющая БЦО, характеризуется состоянием энергообеспеченности, т. е. интенсивностью доставки кислорода и энергосубстратов. Эталоном, пригодным для оценки ФСО, служит базальный (должный) транспорт кислорода (и энергоресурсов), необходимый для поддержания готовности к динамическому самообновлению. Отношение реального транспорта кислорода к эталонному получило название энергодинамического потенциала (ЭДП), нормальный или гипер-, гипо-, а также патоэнергодинамический уровень которого характеризует ФСО.
- Динамическое самообновление является предельной сущностью живой материи. На каждом уровне живой материи — от организменного до биосферного—все без исключения жизненные структуры и процессы являются результатом собственного самообновления, которое обеспечивается обменом энергии и веществ. Саморегулируемое энергообеспечение лежит в основе приспособляемости живых систем к изменяющимся условиям существования и поэтому способных реагировать на внешние и внутренние воздействия динамическим самовосстановлением. Расширение и углубление наших знаний о живом позволяет соотносить систему динамического самовосстановления (СДСВ) с системой энергообеспечения жизнедея-
2 тельности. Только свободная энергия каждой клетки организма обеспечивает физиологическую регенерацию её органелл, репарацию генома, адаптивную стабилизацию внутриклеточных структур и, наконец, репаративную регенерацию (рис. 2.2).
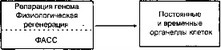
Рис. 2.2. Система динамического самовосстановления (СДСВ)
В организме отсутствует единый энергетический генератор, который централизованно удовлетворял бы все его энергетические потребности. Поэтому задача обеспечения свободной энергией решается каждой клеткой индивидуально. Превращение свободной энергии в элементы СДСВ тесно связано с экспрессией генома (рис. 2.3).
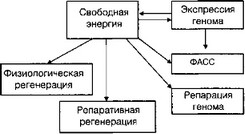
Рис. 2.3. Превращение энергии в процессы СДСВ
Обычно используются три схемы возможного энергетического метаболизма: гликолиз, аэробное окисление субстрата и гексозомонофосфатный шунт. Считается, что наиболее энерге-
тически совершенным, дающим высокий коэффициент свобод- 2
ной энергии на единицу утилизированного субстрата и экологически более чистым является процесс аэробного окисления субстрата. Реализация его локализована в митохондриях — внутриклеточных органеллах, где происходит переработка энергосубстратов в свободную энергию и образуется аденозинтри- фосфат (АТФ), который может использоваться клеткой для выполнения всевозможных энергопотребляющих функций. Клетки сердца, мозга, печени, лёгких, скелетных мышц, почек, особенно нуждающиеся в эффективном энергетическом обеспечении, содержат большое количество митохондрий. Например, до 30% объёма цитоплазмы кардиомиоцитов занимают митохондрии. Энергетически выгодной является реакция гидролиза, в которой молекула воды разрушает ковалентно связанное соединение. Гидролиз одной молекулы АТФ до аденозиндифосфата (АДФ) и неорганического фосфата (Фн) всегда сопровождается выделением так называемой свободной энергии, величина которой при обычных для клетки условиях лежит в пределах от 11 до 13 ккал/моль. Важно, что при отщеплении концевого фосфата исчезает энергетически невыгодное электростатическое отталкивание между соседними отрицательными зарядами. Высвобождающийся при этом неорганический фосфат (ион) стабилизируется благодаря резонансу и образованию энергетически выгодных водородных связей с водой. В случае чередований двойных и простых связей электроны, участвующие в их образовании, перемещаются по молекуле, стабилизируя её структуру, что и называется резонансом. Однако для многих реакций биосинтеза указанного уровня энергии оказывается недостаточно, и тогда путь гидролиза АТФ сначала состоит из образования аденозинмонофосфата (АМФ) и пирофосфата (ПФ), а затем сводится к гидролизу ПФ с образованием свободной энергии в 26 ккал/моль. Во внутриклеточном пространстве типичной
клетки находится около одного миллиарда молекул АТФ, т.е. 109 этих энергетически важных молекул. В таких реакциях роль регулятора обычно выполняет один из ферментов, чаще всего первый, который, как правило, имеет аллостерическую природу. Подобный фермент содержит 2 регуляторных центра, один из которых играет роль позитивного, а другой — негативного эффектора. Субстрат реакции выступает в качестве активатора (позитивного эффектора), а продукт реакции — ингибитора (негативного эффектора). Данная система позволяет поддерживать автоматическую регуляцию механизмов на уровне каждой клетки. Если концентрация АТФ в клетке велика, то ингибируются ферменты, участвующие в его синтезе. При снижении концентрации АТФ ниже нормы и увеличении концентрации АДФ активируется дыхательная цепь, а при росте концентрации АМФ активируется система гликолиза.
При систематически повышенном энергетическом запросе включается более высокий, в данном случае клеточный уровень регуляции энергопреобразующей системы, приводящей к индукции (а при снижении энергетического запроса — к репрессии) синтеза новых ферментов для СДСВ. Индукция или репрессия ферментов является наиболее простым и экономичным способом адаптации клеток к новым условиям самосохранения. Аденилатнуклеотиды, в число которых входят АТФ, АДФ, АМФ и циклический аденозинмонофосфат (цАМФ), являются основными биорегуляторами в СДСВ и воздействуют на различные классы биохимических реакций. В клетках существует ряд и других макроэргов, включая пирофосфат, содержащий две соединённые между собой фосфатные группы (ФФн): креатинин- фосфат, играющий роль энергетического буфера в мышечных клетках, и т.д. Присоединение к различным биохимическим соединениям фосфатсодержащих фрагментов Фн, ФФн, АМФ или АДФ повышает реакционную способность продуктов моди-
фикации и способствует протеканию многих процессов внутри 2 клетки. Последние идут в цитоплазме или матриксе митохондрий. Их относят к реакциям субстратного фосфорилирования, и они протекают только при условии, что это энергетически выгодно. АТФ играет ключевую роль не только в динамическом самосохранении организма, но и в функционировании механизмов удовлетворения всех остальных биологических потребностей его. Принято различать 5 основных энергопотребляющих потоков, связанных с затратами энергии на синтез химических соединений, осмотическую, механическую и электрическую работу, а также продуцирование тепла. АТФ является универсальным преобразователем запасённой энергии в любую из перечисленных выше вщов работ, составляющих компоненты биологического потенциала (БП), как показано на рис. 2.4. Самовосстановление, являясь основой биоусгойчивосги всех клеток, потребляет основные потоки запасаемой энергии. Кроме АТФ, для синтеза белка на рибосомах расходуется гуанозинтрифосфат, в синтезе полисахаридов участвует уридинтрифосфат, а в производстве фосфолипидов — цигазинтрифосфат. Для реализации различных энергетических запросов у клетки имеются надёжные и быстро срабатывающие механизмы по переключению режимов производства свободной энергии с помощью компонентов СДСВ.
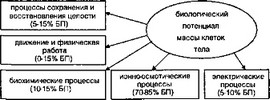
Рис. 2.4. Компоненты биологического потенциала
2 Изменения внутриклеточного содержания АТФ, АДФ, АМФ
и ФФн определяют факт существования нескольких режимов работы митохондрий. В клетках, находящихся в активной фазе деятельности, митохондрии имеют все необходимые ингредиенты для своей работы, включая субстрат для окисления, субстраты для фосфорилирования в виде АДФ и Фн, а также кислород. В этом состоянии митохондрии активно поглощают последний, и процесс окисления сопряжён с фосфорилированием.
В состоянии покоя клеток в митохондриях имеется субстрат для окисления и кислород, но отсутствует АДФ. В этих условиях процесс фосфорилирования становится невозможным, поскольку отсутствует субстрат для фосфорилирования, и митохондрии переходят на более экономный режим работы. При этом скорость дыхания снижается, а концентрация основных переносчиков, в том числе НАДН, растёт. Коэффициент полезного действия митохондрий как энергопреобразующей системы в этом случае приближается к 100%. Принято считать, что в таком состоянии скорость дыхания зависит от логарифма отношения концентраций АТФ к АДФ и Фн. Процесс окисления идёт в равновесных условиях и осуществляется термодинамический контроль реакции. Увеличение энергетического запроса со стороны клетки ведёт к переходу митохондрий в активное состояние. В этом случае термодинамический контроль за работой дыхательной цепи сменяется кинетическим при условии, что кислород не лимитирует перенос электронов по дыхательной цепи. В условиях кинетического контроля баланс в СДСВ зависит от величины энергетического заряда клетки (Qb), определяемого с помощью уравнения:
АТФ+0,5АДФ
Ш=
АТФ+АДФ+АМФ
Регуляторная роль «энергетического заряда» в клетке поддержи- 2
вается балансом всех внутриклеточных метаболических процессов. Обычно величина Qb для клеток в норме лежит в области 0,8— 0,95, что свидетельствует о преобладании в клетке АТФ по сравнению с АДФ и АМФ. Отличительной способностью СДСВ является её абсолютная специфичность: для процессов самовосстановления используется только внутриклеточная свободная энергия и собственная АТФ. Каждый специалист должен знать и помнить, что экзогенная АТФ. вводимая в сосудистое русло, не включается во внушикле- точные энергетические потоки и не участвует в работе СДСВ.
Таким образом, роль СДСВ многогранна и значительна. СДСВ участвует во многих реакциях как регулятор биохимических процессов, в одних случаях активируя их, а в других ингибируя. Особенно велика роль АТФ и его партнёров (АДФ, АМФ, Фн) в регулировании работы дыхательной цепи в зависимости от условий её функционирования. Дисрегуляцию СДСВ вызывает гипоокси- доз, а при отсутствии кислорода СДСВ вообще дезорганизуется.
- Болезнь — нарушение биоустойчивости, вызываемое дисбиотией ЭС и/или дисэнергодинамией ФСО, которое препятствует удовлетворению биологической потребности организма в динамическом самообновлении и сопровождается различными нарушениями обмена веществ и энергообеспечения, ответственными за уменьшение биологической устойчивости.
Дисбиотия ЭС характеризуется отрицательным энергоструктурным балансом, который является следствием нарушений механизмов репарации генома, физиологической регенерации ор- ганелл клеток, феномена адаптивной стабилизации структур и репаративной регенерации.
Дисэнергодинамия ФСО характеризуется энергодефицитом, ответственным за дезинтеграцию активности генов с изменени-
2 ями окружающей среды, диктующими характер экспрессии этих
 генов. ЭС и ФСО находятся в тесном энергопространственном сопряжении, обусловленном биорегуляторными свойствами кислорода при участии воды и ионов. Биорегуляцию осуществляют нервная и гуморальная системы вместе с цитокиновым каскадом (рис. 2.5).
генов. ЭС и ФСО находятся в тесном энергопространственном сопряжении, обусловленном биорегуляторными свойствами кислорода при участии воды и ионов. Биорегуляцию осуществляют нервная и гуморальная системы вместе с цитокиновым каскадом (рис. 2.5).шается динамическое единство ФСО и ЭС, а также сама струк- 2 турно-пространственная упорядоченность компартментов клеток тела, запуская механизмы некробиоза. При адаптивном обеспечении восстановления биоустойчивости структурно-временная дискретность этого процесса слагается из двух последовательных периодов. Первый — подготовительный, продолжительностью минимум 6 часов. Он характеризуется увеличением числа функционирующих структур, активацией транскрипции, трансляции, физиологической регенерации, феноменом адаптивной стабилизации структур. Второй период — суточный, когда, преодолев точку рестрикции, клетки пролиферируют, обеспечивая этим репаративную регенерацию.
Энергоструктурный баланс БЦО представлен четырьмя группами клеток. Первая группа — это клетки, вступившие в апоптоз или подвергнувшиеся некробиозу. Вторую группу составляют гибернирующие и оглушенные клетки. В третью входят дифференцированные клетки, которые и обеспечивают ФСО, благодаря собственной физиологической регенерации. Четвертая группа клеток пролиферирует в ходе репаративной регенерации. Каждую секунду в организме погибает 1 млн. клеток и для сохранения энергоструктурного баланса должно регенерировать такое же количество клеток.
Единицей ЭС является биологический квант (БК). Интенсивность поддержания ЭС отражает удельный биологический квант (УБК). Единицей дискретности ФСО служит энергодинамический квант (ЭДК). Интенсивность проявления ФСО следует оценивать по удельному энергодинамическому кванту (УЭДК). Ритмическая дискретность динамического самосохранения определяется квантовым биоциклом (КБЦ). Коэффициент динамического самосохранения (КДС = реал. КБЩцолж. КБЦ), приближаясь к 1, отражает высокую потенциальную возможность системы динамического самовосстановления. Коэффициент сопряжен-
2 ности УЭДК и УБК характеризует минимальную функциональ
но-структурную дискретность (КС = реал. КС/долж. КС). В отсутствие энергодефицита КСgt;1 характеризует установившееся структурно-функциональное равновесие. Особенности нестабильности БЦО определяют гомеокинетические свойства КБЦ.
Система восстановления локальной целости осуществляет тканевый морфогенез путем реакции воспаления, обеспечивая самовозобновление тканей и органов.
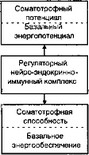
Система восстановления биоустойчивости реализуется через общий адаптационный синдром (ОАС), который организуется ЦНС через полигландулярную соматотрофную интенсификацию. В запуске ОАС большую роль играет ноцицептивный эквивалент биоэнергетической гипоксии. Биологическим смыслом адаптации является сохранение БЦО (рис. 2.6).
Слабые звенья первыми реагируют на любой стрессор и поэтому ответственны за любую дезадаптацию, являющуюся результатом отрицательного энергоструктурного баланса, вызываемого некрозом, апоптозом, гибернацией и оглушенностью клеток функциональных систем организма. Возникшие повреждения устраняются системой восстановления БЦО в ходе ликвидации энергоструктурного дефицита. Продолжительность уравновешивания структурных повреждений развертыванием ОАС может быть минимизирована 54 часами: 6 часов адаптационной стабилизации внутриклеточных структур для подготовки к регенерации и 48-часовой оптимальный период 2-х клеточных циклов. Таким образом, стадия резистентности ОАС, обеспечивающая энергоструктурный баланс, может быть достигнута на 3-и пос- тагрессивные сутки, если будут использоваться возможности ИМ. Её эффективность прежде всего зависит от информативности специального мониторинга, который был назван статуе- метрией. Это биологический мониторинг, базирующийся на непрерывном количественном определении угрозоопаснос
ти дефицита энергодинамического потенциала (нарушений ФСО) и критичности утраты биологического потенциала (нарушений ЭС). Путем изучения угрозоопасных и критических нарушений непрерывно определяется интегральный параметр сга- тусметрии — клинический индекс тяжести статуса (КИТС). Он объединяет угрозоопасность с критичностью и поэтому очень точно определяет риск нарушений биоустойчивости.