
Орган зрения состоит из глазного яблока, соединённого через зрительный нерв с мозгом, и включает вспомогательный аппарат в виде век, слёзной железы и поперечнополосатых глазодвигательных мышц.
- Развитие (рис. 8-43 и 8-44). Зачаток глаза появляется у 22-дневного эмбриона как пара неглубоких желобков в выростах переднего мозга. После закрытия нейропоров эти выросты образуют глазные пузыри.
а. Глазные пузыри связаны с эмбриональным мозгом при помощи глазных стебельков. Глазные пузыри вступают в контакт с эктодермой будущей лицевой части головы и индуцируют в ней развитие хрусталика. Инвагинация стенки глазного пузыря приводит к формированию двухслойного глазного бокала.
б. Глазной бокал
- Наружный слой глазного бокала образует пигментный слой сетчатки.
- Внутренний слой формирует сетчатку. Аксоны дифференцирующихся ганглиозных клеток прорастают в глазной стебелёк и входят в состав зрительного нерва.
в. Сосудистая оболочка формируется из окружающей глазной бокал мезенхимы.

Рис. 8-43. Развитие глаза. А — 22-дневный эмбрион; Б — 4-недельный эмбрион; В — эмбрион длиной 5 мм [из Sadler TW, 1990]
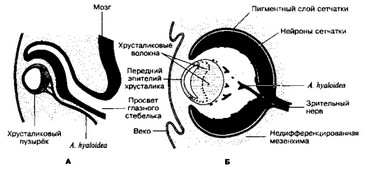
Рис. 8-44. Развитие глаза. А — 6-недельный зародыш; Б — 7-недельный зародыш [из Mann IC, 1974]
Эктомезенхима. Особое значение имеют выселяющиеся из нервного гребня клетки т.н. эктомезенхимы, участвующие в образовании склеры и цилиарной мышцы, а также дифференцирующиеся в эндотелиальные клетки и фибробласты роговицы.
г. Эпителий роговицы развивается из эктодермы.
д. Хрусталик. Зачаток хрусталика отделяется от эктодермы и превращается в хрусталиковый пузырёк, над которым смыкается эктодерма. При развитии хрусталикового пузырька изменяется толщина его стенок, в связи с чем появляются более тонкий передний эпителий и комплекс плотно упакованных хрусталиковых волокон на задней поверхности хрусталикового пузырька. Хрусталиковые волокна удлиняются и заполняют полость пузырька. В эпителиальных клетках хрусталика синтезируются специфические для хрусталика белки — кристаллины. На начальных стадиях дифференцировки клетки хрусталика синтезируют небольшое количество а- и р-кристал- линов. По мере дифференцировки удлинённые клетки хрусталика, кроме а- и р-крис- таллинов, начинают синтезировать у-кристаллины.
- Глазное яблоко. Стенка глазного яблока образована оболочками: в передней части — роговица, в задней — сетчатка, сосудистая оболочка и склера (рис. 8-58).
а. Роговица — прозрачная оболочка передней стенки глаза, состоит из пяти слоёв. Лимб — граница между прозрачной роговицей и непрозрачной склерой.
Слои роговицы (рис. 8-58)
(а) Многослойный плоский неороговевающий эпителий.
(б) Передняя пограничная мембрана (боуменова оболочка) — не содержа
щий клеток гомогенный слой основного вещества и неупорядоченно ориентированных тонких коллагеновых и ретикулиновых волокон; поддерживает форму роговицы.
(в) Собственное вещество представлено правильно расположенными коллагеновыми пластинками и уплощёнными фибробластами.
(г) Задняя пограничная мембрана (десцеметова оболочка) — прозрачный слой роговицы, расположен между собственным веществом и эндотелием задней поверхности роговицы, состоит из коллагеновых волокон (коллаген типа VID) и аморфного вещества.
(д) Эндотелий ограничивает спереди переднюю камеру глаза.
б. Склера — наружная непрозрачная оболочка глазного яблока. Склера построена из плотных тяжей коллагеновых волокон, между которыми находятся уплощённой формы фибробласты. В месте соединения склеры с роговицей расположены небольшие сообщающиеся полости, в совокупности образующие шлёммов канал, обеспечивающий отток жидкости из передней камеры глаза.
в. Сосудистая оболочка осуществляет питание сетчатки. Эта оболочка состоит из нескольких слоёв (пластинок). Радужная оболочка — передний вырост сосудистой оболочки. Другая часть сосудистой оболочки, цилиарное тело, участвует в аккомодации зрения, регулируя форму хрусталика.
- Пластинки
(а) Надсосудистая. Расположена на границе со склерой, состоит из рыхлой волокнистой соединительной ткани.
(б) Сосудистая. Содержит сплетение артерий и вен. В рыхлой соединительной ткани располагаются пигментные клетки и ГМК.
(в) Хориокапиллярная. Образована сплетением капилляров синусоидного типа.
(г) На границе с сетчаткой располагается базальная пластинка.
- Радужная оболочка — продолжение сосудистой оболочки глаза, расположена между роговицей и хрусталиком, разделяет переднюю и заднюю камеры глаза.
(а) Слои (от передней к задней камере): эндотелий, наружный пограничный, сосудистый, внутренний пограничный, пигментный.
(б) Мышцы. В состав радужки входят суживающая (циркулярная) и расширяющая зрачок мышцы. При раздражении парасимпатических (холинерги- ческих) нервных волокон зрачок суживается, симпатическая стимуляция приводит к расширению зрачка.
(в) Цвет глаз определяют количество и тип пигмента в радужной оболочке. Например, в глазах голубого цвета мало меланоцитов и соответственно пигмента. Альбинизм (см. главу 18).
- Цилиарное тело. В области угла глаза сосудистая оболочка утолщается, образуя цилиарное тело, имеющее на срезе вид треугольника, обращённого основанием в переднюю камеру. Основную массу цилиарного тела занимает цилиарная мышца, играющая важную роль в аккомодации глаза. В её составе ГМК проходят в трёх взаимно перпендикулярных направлениях. От цилиарного тела отходят по направлению к хрусталику цилиарные отростки, к которым прикрепляется цйннова связка. При сокращении цилиарной мышцы циннова связка расслабляется, и выпуклость хрусталика увеличивается.
г. Хрусталик имеет вид двояковыпуклого тела. Его передняя стенка состоит из однослойного кубического эпителия, который по направлению к экватору становится выше. Эпителиальные клетки хрусталика связаны щелевыми контактами. Удлинённые веретенообразной формы эпителиальные клетки, достигшие состояния терминальной дифференцировки, содержат кристаллины и с возрастом утрачивают ядра и органеллы. Это прозрачные хрусталиковые волокна, составляющие основную часть хрусталика. Капсула хрусталика — толстая базальная мембрана. Прозрачность хрусталика и/или его капсулы нарушается при катаракте.
д. Стекловидное тело — прозрачная среда глаза, заполняет полость между хрусталиком и сетчатой оболочкой; стекловидное тело — гель, содержащий воду, коллаген, белок витреин и гиалуроновую кислоту. Через стекловидное тело от сетчатки к хрусталику проходит канал — остаток эмбриональной сосудистой системы глаза (рис. 8-44).
е. Сетчатая оболочка (сетчатка) — внутренняя оболочка глаза, имеет зрительный отдел, по зубчатому краю переходящий в слепой отдел, покрывающий сзади цилиарное тело и радужку. У заднего края оптической оси глаза сетчатка имеет округлое жёлтое пятно диаметром около 2 мм. Центральная ямка — углубление в средней части жёлтого пятна, место наилучшего восприятия. Зрительный нерв выходит из сетчатки медиальнее жёлтого пятна. Здесь образуется диск зрительного нерва (слепое пятно). В центре диска имеется углубление, в котором видны питающие сетчатку сосуды, выходящие из зрительного нерва.
- Слои
(а) Пигментный. Клетки полигональной формы, прилежащие к сосудистой оболочке. Одна пигментная клетка взаимодействует с наружными сегментами десятков фоторецепторных клеток — палочек и колбочек (рис. 8-45).
(б) Наружный ядерный. Ядросодержащие части фоторецепторных клеток. Колбочки концентрируются в области жёлтого пятна. Глазное яблоко организовано таким образом, что на колбочки падает центральная часть светового пятна от визуализируемого объекта. По периферии от жёлтого пятна расположены палочки.
(в) Наружный сетчатый. Здесь осуществляются контакты внутренних сегментов палочек и колбочек с дендритами биполярных клеток.
(г) Внутренний ядерный. Здесь располагаются биполярные клетки, связывающие палочки и колбочки с ганглиозными клетками, а также горизонтальные и амакринные клетки. Перикарионы амакринных клеток расположены во внутренней части внутреннего ядерного слоя.
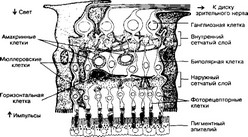
Рис. 8-45. Сетчатка. Стрелками слева обозначены направления светового потока и возбуждения. Пигментный эпителий окружает наружные сегменты фоторецепторных клеток, образующих синаптические контакты с биполярными нейронами. Информация от биполярных клеток передаётся ганглиозным клеткам и по их аксонам, образующим зрительный нерв, уходит в мозг. Промежутки между нейронами заполняют крупные мюллеровские клетки. Их наружные отростки заканчиваются на границе между наружными и внутренними сегментами фоторецепторных клеток [из Boycott, Dowling, 1966]
(д) Внутренний сетчатый. В нём биполярные клетки контактируют с ганглиозными клетками, амакринные клетки выступают в качестве вставочных нейронов. Популярна концепция о том, что ограниченное число биполярных клеток передает информацию 16 типам ганглиозных клеток при участии не менее 20 типов амакринных клеток.
(е) Ганглиозный слой содержит ганглиозные нейроны. Общая схема передачи информации в сетчатке такова: рецепторная клетка -gt; биполярная клетка -gt; ганглиозная клетка и одновременно амакринная клетка -gt; ганглиозная клетка.
- Фоторецепторные клетки — палочки (рис. 8-46) и колбочки. Различают центральное и периферическое зрение, что связано с характером распределения в сетчатке палочек и колбочек. Периферические отростки фоторецепторных клеток состоят из наружного и внутреннего сегментов, соединённых ресничкой.
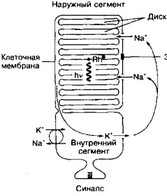
(а) Наружный сегмент имеет множество уплощённых замкнутых дисков, содержащих зрительные пигменты: родопсин — в палочках; красный, зелёный и синий пигменты — в колбочках.
(б) Внутренний сегмент заполнен митохондриями и содержит базальное тельце, от которого в наружный сегмент отходит 9 пар микротрубочек.
(в) Центральное зрение, а также острота зрения реализуются колбочками.
(д) Периферическое зрение, а также ночное зрение и восприятие подвижных объектов — функции палочек.
(е) Цветовосприятие — функция колбочек. Существует три типа колбочек, каждый из которых содержит только один из трёх разных (красный, зелёный и синий) зрительных пигментов.
- Зрительный пигмент состоит из апопротеина (опсин), ковалентно связанного с хромофором (11-цис-ретиналь или 11-цис-дегидроретиналь).
-
 Спектральная чувствительность красного, зелёного и синего зрительных пигментов различна — соответственно 560, 535 и 440 нм — и определяется первичной структурой апопротеина.
Спектральная чувствительность красного, зелёного и синего зрительных пигментов различна — соответственно 560, 535 и 440 нм — и определяется первичной структурой апопротеина.
Рис. 8-46. Схема палочки. В наружном сегменте расположена стопка дисков, содержащих зрительный пигмент родопсин. Мембрана дисков и клеточная мембрана разобщены. Свет (hv) активирует родопсин Rh* в дисках, что закрывает Ыа*-каналы в клеточной мембране и снижает вход Na+ в клетку [из Lamb TD, 1986]
- Трихромазйя — возможность различать любые цвета, определяется присутствием в сетчатке всех трёх зрительных пигментов (для красного, зелёного и синего — первичные цвета). Эти основы теории цветного зрения предложил Томас Янг (1802).
- Дихромазйи — дефекты цветового восприятия (преимущественно у мужчин; например, в Европе разные дефекты у мужчин составляют 8% общей популяции) по одному из первичных цветов — подразделяют на протанопии, дейтано- пии и тританопии {от гр. первый, второй и третий [имеются в виду порядковые номера первичных цветов: соответственно красный, зелёный, синий])
- Протанопия (страдает восприятие красного, примерно 25% случаев цветовой слепоты) развивается при связанном с хромосомой X наследовании генного дефекта.
- Дейтанопия (цветовая слепота по восприятию зелёного, около 75% всех случаев; связанное с хромосомой X наследование, полиморфизм гена).
- Тританопия (страдает преимущественно восприятие фиолетового цвета, дефектное зрение по синему и жёлтому). Аутосомное доминантное наследование дефектного гена (7q31.3-q32),
- Нейроны
(а) Горизонтальные клетки. Их перикарионы расположены в наружной части внутреннего ядерного слоя, а отростки входят в область синапсов между фоторецепторными и биполярными клетками. Горизонтальные клетки получают информацию от колбочек и передают её также колбочкам. Соседние горизонтальные клетки связаны между собой щелевыми контактами.
(б) Амакринные клетки. Их перикарионы находятся во внутренней части внутреннего ядерного слоя в области синапсов между биполярными и ганглиозными клетками.
(в) Ганглиозные клетки — крупные мультиполярные нейроны многих разновидностей. Их аксоны образуют зрительный нерв.
- Нейромедиаторы. Нейроны сетчатки синтезируют ацетилхолин, дофамин, L-глутаминовую кислоту, глицин, у-аминомасляную кислоту. Некоторые нейроны содержат серотонин, его аналоги (индоламины) и нейропептиды.
- Функциональные особенности
(а) Биполярные клетки реагируют на контрастность изображения. Некоторые биполяры сильнее реагируют на цветной, нежели на чёрно-белый контраст. Одни получают информацию преимущественно от палочек, другие — от колбочек.
(б) Ганглиозные клетки реагируют на множество свойств зрительного объекта (например, на светлые и тёмные объекты, однородность освещения, цвет объекта, его ориентацию).
- Центральная ямка. В области центральной ямки расположены преимущественно колбочки. Каждая колбочка центральной ямки образует синапс только с одним биполярным нейроном. Внутренний ядерный и ганглиозный слои в области центральной ямки истончены.
- Глия. Кроме нейронов, сетчатка содержит крупные клетки радиальной глии — мюмеровские клетки (рис. 8-45). Их ядра расположены на уровне центральной части внутреннего ядерного слоя. Наружные отростки заканчиваются микроворсинками, образуя наружный пограничный слой. Внутренние отростки имеют расширение (ножку) во внутреннем пограничном слое на границе со стекловидным телом.
 Функция. Глиальные клетки играют важную роль в регуляции ионного гомеостаза сетчатки. В частности, они снижают концентрацию K+ во внеклеточном
Функция. Глиальные клетки играют важную роль в регуляции ионного гомеостаза сетчатки. В частности, они снижают концентрацию K+ во внеклеточномпространстве, где концентрация этих ионов при световом раздражении резко увеличивается. Плазматическая мембрана мюллеровских клеток в области ножки характеризуется высокой проницаемостью для ионов K+, выходящих из клетки. Мюллеровская клетка захватывает K+ из наружных слоёв сетчатки и направляет поток этих ионов через свою ножку в жидкость стекловидного тела.
- Механизм фотовосприятия. В состав дисков фоторецепторных клеток входят зрительные пигменты, в т.ч. родопсин палочек.
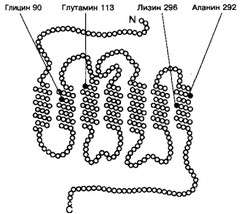
(а) Родопсин (рис. 8-47) состоит из белковой части (опсин) и хромофора — 11 -цис- ретиналя, под действием фотонов переходящего в /и/иис-ретиналь. Мутации генов, кодирующих синтез опсинов, приводят к развитию пигментного ретинита и ночной (куриной) слепоты. Описано около 40 мутаций генов опсинов.
(б) Каскад фотоактивации (рис. 8-48). При попадании квантов света на наружные сегменты в фоторецепторных клетках последовательно происходят следующие события: активация родопсина в результате фотоизомеризации -gt; каталитическая активация G-белка (G1, трансдуцин) [глава 2 I В 2 а (3) (a) (i)]
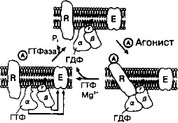 Рис. 8-48. Трансмембранный белок родопсин и его связь с G-белком (трансдуцин) в плаз- молемме фоторецепторных клеток. Возбуждённый фотонами родопсин активирует G-белок. При этом гуанозиндифосфат, связанный с а-СЕ G-белка, заменяется на гуанозинтрифосфат. Отщеплённые а-СЕ и P-CE действуют на фосфоди- эстеразу и заставляют её превращать цГМФ в гуанозинмонофосфат. Это закрывает Ыа’-каналы, и ионы Na+ не могут попасть в клетку, что приводит к её гиперполяризации. R — родопсин; а, р и у — CE G-белка; А — агонист (в данном случае кванты света); E — фермент-эффектор фосфоди- эстераза [из Dratz EA etal, 1993]
Рис. 8-48. Трансмембранный белок родопсин и его связь с G-белком (трансдуцин) в плаз- молемме фоторецепторных клеток. Возбуждённый фотонами родопсин активирует G-белок. При этом гуанозиндифосфат, связанный с а-СЕ G-белка, заменяется на гуанозинтрифосфат. Отщеплённые а-СЕ и P-CE действуют на фосфоди- эстеразу и заставляют её превращать цГМФ в гуанозинмонофосфат. Это закрывает Ыа’-каналы, и ионы Na+ не могут попасть в клетку, что приводит к её гиперполяризации. R — родопсин; а, р и у — CE G-белка; А — агонист (в данном случае кванты света); E — фермент-эффектор фосфоди- эстераза [из Dratz EA etal, 1993]
родопсином —gt; активация фосфодиэстеразы при связывании с G(a —gt; гидролиз цГМФ цГМФ-фосфодиэстеразой -gt; переход цГМФ-зависимых Ыа+-каналов из открытого состояния в закрытое -gt; гиперполяризация плазмолеммы фоторецепторной клетки -gt; передача сигнала на биполярные клетки.
- Активация родопсина в результате фотоизомеризации. Свет, поглощаемый родопсином, инициирует ответ в каскаде цГМФ.
- Каталитическая активация G-белка (Gt, трансдуцин) родопсином. Активированный родопсин взаимодействует с G-белком (состоит из трёх CE), а- CE G-белка активирует цГМФ-фосфодиэстеразу.
- Активация фосфодиэстеразы при связывании с G,а. Увеличение активности цГМФ-фосфодиэстеразы снижает концентрацию цГМФ, что сопровождается закрытием ионных каналов.
- Гиперполяризация плазмолеммы фоторецепторной клетки — следствие закрытия ионных каналов. В результате рецепторная клетка гиперполя- ризуется, что служит сигналом для изменения характера секреции медиатора в синапсе между внутренним сегментом рецепторной клетки и дендритом биполярной клетки. Если рецепторные клетки отвечают на свет гиперполяризацией, то другие типы нейронов сетчатки при этом гиперполяризуются или деполяризуются.
(в) Темновой ток. В темноте ионные каналы в клеточной мембране рецепторных клеток поддерживаются в открытом состоянии за счёт связывания белков ионных каналов с цГМФ. Потоки внутрь клетки Na+ и Ca2* через открытые каналы обеспечивают темновой ток.