Адаптивный иммунный ответ
Первая линия защиты должна обеспечивать элиминацию патогена. Однако это происходит не всегда. В таких случаях запускается вторая линия защиты, связанная с развитием адаптивного иммунного ответа, основные типы которого представлены на рис. 1.5, а стадии развития — на рис. 1.6. Развитие иммунного ответа включает индуктивную и продуктивную (эффекторную) фазы. Индуктивная фаза состоит в формировании исполнительных механизмов адаптивного иммунитета и реализуется в первые 7 сут после появления патогена в организме. В этот период основную роль в иммунной защите играет врожденный иммунитет. По мере развития эффекторных механизмов адаптивного иммунитета они берут на себя основную нагрузку в защите организма.
Одновременно с активацией факторов врожденного иммунитета происходит событие, иллюстрирующее связь между врожденным и адаптивным иммунитетом. Дендритные клетки, как и макрофаги, присутствующие в барьерных тканях, поглощают патогены или их фрагменты и транспортируют их в региональный лимфатический узел. В процессе перемещения в гранулах этих клеток происходит расщепление поглощенных антигенов на фрагменты, взаимодействующие с молекулами MHC-II, и транспорт образовавшихся комплексов на клеточную поверхность. Это необходимо для запуска адаптивного иммунитета, поскольку Т-лимфоциты способны распознавать антиген только в комплексе с молекулой MHC. Таким образом, дендритная клетка вовлекает в реакцию Т-лимфоциты, ответственные за запуск других клеток адаптивного иммунитета. В лимфатических узлах происходит взаимодействие дендритных клеток с Т-лимфоцитами, распознающими антигенные пептиды в составе молекул MHC на мембране дендритной клетки. Это взаимодействие облегчается локализацией дендритных клеток и Т-лимфоцитов в одной и той же зоне лимфатического узла (Т-зоне), обусловленной влиянием одних и тех же хемотаксических
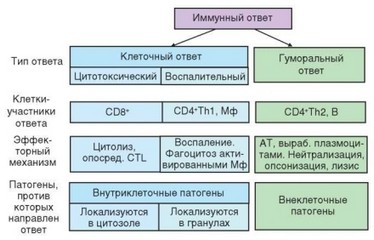
Рис. 1.5. Типы иммунного ответа, развивающегося в ответ на действие патогенов с различной локализацией (участвующие клетки и механизмы)
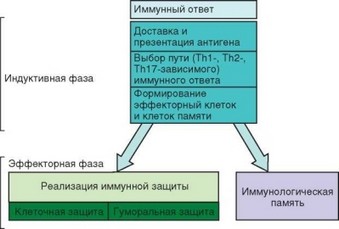
Рис. 1.6. Основные события иммунного ответа
сигналов. Дендритные клетки проникают в Т-зоны лимфогенным, а рецир- кулирущие Т-лимфоциты — гематогенным путем.
TCR распознает антигенный пептид в составе молекул MHC с участием корецепторов CD4 (Т-хелперы) и CD8 (цитоткосические Т-клетки или Т-киллеры). Этому способствует сродство корецепторов к молекулам MHC (CD4 — к MHC-II, CD8 — к MHC-I). Процесс представления антигенного пептида Т-клеткам в составе молекул MHC называют презентацией антигена. Таким образом, Т-хелперы распознают комплекс MHC-П-антигенный пептид. Это служит необходимым, но недостаточным условием для активации Т-клеток. Дополнительные активационные стимулы клетки получают через костимулирующие молекулы. В их роли на дендритных клетках выступают молекулы В7 (CD80 и CD86), а на Т-клетках — CD28. Сигнал, поступающий в Т-клетку через молекулу CD28, дополняет сигнал от TCR, что обеспечивает активацию клетки. Если CD28 исходно присутствует на большинстве Т-клеток, то молекула CD80 на дендритных клетках барьерных тканей почти отсутствует, а CD86 представлена на них в количествах, недостаточных для костимуляции. Усиление экспрессии молекул В7 на дендритных клетках (а также макрофагах и других клетках, способных пре- зентировать антиген) происходит при распознавании PAMP и тесно связано с другими проявлениями врожденного иммунитета. Таким образом, для запуска адаптивного иммунитета необходима предварительная активация клеток врожденного иммунитета.
При активации Т-клетки начинают экспрессировать гены своего ростового фактора — IL-2 и его рецептора. В результате аутокринного действия IL-2 Т-клетки активированных клонов пролиферируют. Пролиферции, индуцированной антигеном, подвергаются также Т-киллеры и В-лимфо- циты. Поскольку исходно число клеток каждого клона невелико, пролиферация имеет большое значение для обеспечения эффективной иммунной защиты (7—8 делений обеспечивают увеличение числа клеток в 250—500 раз). На следующем этапе CD4+ Т-клетки дифференцируются на основные разновидности Т-хелперов, из которых упоминаются наиболее изученные — Th1- и ТИ2-клетки. Эти клетки различаются, главным образом, спектром продуцируемых цитокинов, отвечающих за развитие двух важных ветвей иммунного ответа — клеточного, направленного на элиминацию внутриклеточных патогенов, и гуморального, играющего основную роль в борьбе с внеклеточными патогенами и макропаразитами.
Окончательная дифференцировка Т-хелперов происходит в Т-зонах вторичных лимфоидных органов. Thl-клетки мигрируют из Т-зон в очаги воспаления, в том числе в места проникновения патогенов, вызывающих иммунный ответ. Распределение активированных и эффекторных Т-лимфо- цитов отличается от такового наивных Т-клеток, что обусловлено изменением набора экспрессируемых молекул адгезии («рецепторов хоминга») и хемоки- новых рецепторов. Эффекторные Т-лимфоциты мигрируют преимущественно в те отделы иммунной системы, из которых дендритные клетки доставили антиген в лимфатический узел. Thl-клетки реализуют свою активность, кооперируя с макрофагами, а ^2-клетки — с В-лимфоцитами.
Запуск Thl-зависимого иммунного ответа происходит, если клетка фагоцитировала микроорганизм, но не смогла его убить и расщепить. В этом случае фагоцит (макрофаг) получает стимулирующие сигналы со стороны Т-хелперов типа Thl. Взаимодействие между клетками осуществляется в форме повторной презентации антигенного пептида Т-клетке — на этот раз макрофагом. В виде ответной реакции Thl-клетка активируют макрофаг, передавая костимулирующий сигнал через мембранную молекулу CD40 и секретируемый цитокин — интерферон (IFN) у.
В-клетка распознает антиген в его нативном состоянии без участия дендритной клетки. Более того, она сама выступает в роли АПК: поглотив комплекс антигена с рецептором, В-лимфоцит обрабатывает его, встраивая антигенный пептид в состав молекулы MHC-II. В-клетка презентирует пептид ^2-клетке, получая при этом от нее активационные сигналы через ту же костимулирующую молекулу CD40 (как и макрофаги) и цитокин IL-4 (фактор роста В-клеток). Это вызывает сначала пролиферацию клонов активированных В-лимфоцитов (в реакцию вовлекаются клоны, распознавшие антиген), а затем их дифференцировку в антителообразующие клетки — плазмоциты. Эти процессы проходят наиболее эффективно в зародышевых центрах, формирующихся при иммунном ответе в лимфоидных фолликулах (при этом происходит превращение первичных фолликулов во вторичные).
В зародышевых центрах В-клетки интенсивно пролиферируют, при этом происходит резкое (на 4—6 порядков) повышение частоты соматических мутаций в гипервариабельных зонах вариабельных генов иммуноглобулинов. В клетках ослабляется экспрессия факторов, предотвращающих развитие апоптоза (программированной гибели клеток), что приводит к быстрой гибели В-лимфоцитов при отсутствии дополнительных сигналов к выживанию. Таким сигналами служит распознавание рецепторами BCR антигенов в составе иммунных комплексов, которые локализуются на поверхности фолликулярных дендритных клеток. Ограниченное содержание антигена в зародышевых центрах вызывает конкуренцию за его связывание между антигенспецифичными В-клетками, в которой побеждают В-клетки, несущие BCR, обладающие наибольшим сродством к антигену. Этот процесс называют созреванием аффинитета антигенраспознающих рецепторов (или антител, поскольку мембранные рецепторы на следующих этапах дифференцировки В-клеток будут секретироваться в виде растворимых иммуноглобулинов — антител). Параллельно происходит переключение изотипа секретируемых иммуноглобулинов с IgM на IgG, IgA и IgE. Процесс завершается в апикальной части зародышевых центров превращением активированных В-клеток (В-лимфобластов) в плазматические клетки, непосредственно секретирующие антитела. Одновременно с образованием плазматических клеток В-лимфоциты дифференцируются в В-клетки памяти. Плазматические клетки мигрируют в другие отделы иммунной системы (маргинальную зону селезенки, мозговые шнуры лимфатических узлов и т.д.). Большинство их попадает в костный мозг. Микроокружение этого органа обеспечивает длительное существование плазмоцитов.
Особую роль играет третий вариант реакции адаптивного иммунитета — ответ цитотоксических Т-лимфоцитов. Презентацию антигенного пептида этим Т-клеткам тоже осуществляют дендритные клетки, но с участием молекул MHC-I. Цитотоксические клетки, предназначенные для защиты от вирусов и других внутриклеточных патогенов (присутствуют в цитозоле), в меньшей степени зависят от Т-хелперов. Контактные взаимодействия цитотоксических Т-клеток с Т-хелперами не играют такой важной роли, как для эффекторных клеток при развитии двух других рассмотренных выше форм ответа. Акцессорная функция Th1-клеток в реакциях цитотоксичности состоит, главным образом, в снабжении Т-киллеров необходимым для экспансии их клонов ростовым цитокином IL-2. Цитотоксические Т-лимфо- циты развиваются в Т-зонах лимфоидных органов, а затем расселяются по организму, в основном мигрируя в очаги воспаления и барьерные ткани.
В результате описанных выше процессов происходит активация клеток сначала врожденного, а затем адаптивного иммунитета, причем связующим звеном между ними служат дендритные клетки. При этом происходит активация всех клеток врожденного иммунитета в очаге проникновения патогена и развития воспаления, тогда как в системе адаптивного иммунитета в активацию вовлекаются только специфичные к распознаваемым антигенам клоны лимфоцитов. При этом cуществует определенная избирательность в вовлечении различных факторов врожденного и адаптивного иммунитета в зависимости от природы возбудителя (бактерии или вирусы, внеклеточные или внутриклеточные патогены и т.д.).
Таким образом, запуск адаптивного иммунного ответа невозможен без участия факторов врожденного иммунитета, прежде всего активированных дендритных клеток, презентирующих антиген Т-клеткам и экспрессирующих костимулирующие молекулы. Антиген только отбирает клоны лимфоцитов, которые будут активированы. Для активации клеток, распознавших антиген, они должны получить дополнительные сигналы через костиму- лирующие молекулы. При этом активационную «эстафету» от дендритных клеток принимают CD4+ Т-хелперы, которые расширяют спектр вовлекаемых в иммунный ответ клеток, оказывая костимулирующее действие на В-лимфоциты, макрофаги и снабжая ростовым фактором цитотоксические
Т-лимфоциты. Подобная схема развития иммунного ответа соответствует ситуации, когда антиген действует совместно с PAMP, что имеет место при инфицировании — попадании в организм потенциально патогенных микроорганизмов.
Однако часто антиген действует на лимфоциты, в том числе Т-клетки, изолированно от PAMP. Это бывает при приеме пищи, росте иммуногенной опухоли, при пересадке чужеродных органов, действии аллергенов. В отношении пищевых антигенов развивается анергия, т.е. активная форма «неотвечаемости», распространяющаяся только на данный антиген. Вероятно, развитие анергии происходит и при росте опухоли, однако при этом действует другой сигнал опасности — экспрессия стрессорных молекул, активирующих NK-клетки. Природа дополнительного сигнала, участвующего в развитии аллергического ответа в отсутствие PAMP, не ясна. Вероятно, именно особенности этого сигнала определяют выбор ^2-типа иммунного ответа на аллергены. Как отмечалось выше, в норме иммунная система не реагирует на собственные антигены организма (аутоантигены). Однако при локальном воспалении, сопровождающемся экспрессией кости- мулирующих молекул на соматических клетках (не дендритных), а также в некоторых других ситуациях может развиваться аутоиммунная патология.
Источник: Ярилин.А.А , «Иммунология » 2010
А так же в разделе «Адаптивный иммунный ответ »
- Зарождение иммунологии
- Развитие иммунологии до середины ХХ века
- «Новая иммунология» 50-80-х годов ХХ века
- Современный этап развития иммунологии — молекулярная иммунология
- ЕСТЕСТВЕННАЯ ИСТОРИЯ ИММУНИТЕТА
- КРАТКОЕ ИЗЛОЖЕНИЕ ИММУНОЛОГИИ
- Молекулы-мишени иммунитета (образы патогенности, антигены) и распознающие их рецепторы
- Иммунная система
- Первая линия иммунной защиты
- Эффекторные механизмы иммунного ответа. Взаимосвязь факторов врожденного и адаптивного иммунитета
- Иммунологическая память
- РЕЗЮМЕ